Plik kwas fosfatydowy lub fosfatydat, To fosfolipid należący do rodziny glicerofosfolipidów lub fosfoglicerydów, które są obecne we wszystkich błonach biologicznych. Jest to najprostszy fosfolipid i działa jako prekursor innych, bardziej złożonych glicerofosfolipidów, chociaż nie występuje w dużych ilościach.
Na E coli, na przykład stanowi mniej niż 0,5% wszystkich fosfolipidów w błonie komórkowej i zmienia się szybko ze względu na swoją rolę jako biosyntetyczny związek pośredni.

Ten prekursorowy fosfolipid jest tworzony przez acylowanie grup hydroksylowych 3-fosforanu glicerolu dwoma aktywowanymi cząsteczkami kwasu tłuszczowego i uważa się, że jest obecny w praktycznie wszystkich błonach biologicznych..
Kardiolipina, ważny fosfolipid obecny w błonie mitochondrialnej i błonie plazmatycznej bakterii i archeonów, składa się z dwóch cząsteczek kwasu fosfatydowego przyłączonych do cząsteczki glicerolu.
Kwas lizofosfatydowy, czyli cząsteczka kwasu fosfatydowego pozbawiona grupy acylowej, uczestniczy jako cząsteczka pośrednia w wielu procesach sygnalizacji pozakomórkowej.
Indeks artykułów
Podobnie jak większość fosfolipidów, kwas fosfatydowy jest amfipatyczną cząsteczką z dwoma końcami o przeciwnej hydrofilowości: hydrofilowym końcem polarnym i hydrofobowym niepolarnym ogonem..
Jak wspomniano wcześniej, jest to najprostszy fosfolipid, ponieważ jego „głowa” lub grupa polarna składa się tylko z grupy fosforanowej, która jest przyłączona do węgla w pozycji 3 cząsteczki glicerolu..
Ich apolarne ogony składają się z dwóch łańcuchów kwasów tłuszczowych zestryfikowanych z atomami węgla w pozycjach 1 i 2 3-fosforanu glicerolu. Te kwasy tłuszczowe mają różne długości i stopnie nasycenia.
Zwykle długość przyłączonych kwasów tłuszczowych wynosi od 16 do 24 atomów węgla; i ustalono, że kwas tłuszczowy przyłączony do węgla 2 jest zwykle nienasycony (obecność podwójnych wiązań węgiel-węgiel), chociaż zależy to od rozpatrywanego organizmu, ponieważ w plastydach roślin jest to nasycony kwas tłuszczowy.
Biosynteza kwasu fosfatydowego jest punktem rozgałęzienia syntezy innych glicerofosfolipidów. Rozpoczyna się aktywacją kwasów tłuszczowych poprzez dodanie części CoA, reakcję katalizowaną przez syntetazę acylo-CoA, która wytwarza acylo-CoA.
Istnieją różne izoformy tego enzymu występujące w siateczce endoplazmatycznej i mitochondriach, ale reakcje zachodzą w podobny sposób, jak u prokariotów..
Pierwszym „zaangażowanym” etapem szlaku biosyntezy jest przeniesienie cząsteczki acylo-CoA do 3-fosforanu glicerolu, reakcja katalizowana przez acylotransferazę glicerolu 3-fosforanu związaną z zewnętrzną błoną mitochondriów i siateczką śródplazmatyczną..
Uważa się, że produkt tej reakcji, kwas lizofosfatydowy (ponieważ ma tylko jeden łańcuch węglowodorowy), jest przenoszony z mitochondriów do retikulum endoplazmatycznego w celu przeprowadzenia drugiej reakcji acylowania..
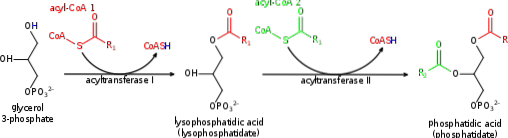
Enzym katalizujący ten etap jest znany jako acylotransferaza 1-acyloglicerolo-3-fosforanu, występująca w błonie retikulum endoplazmatycznego i który specyficznie przenosi nienasycone kwasy tłuszczowe do węgla w pozycji 2 cząsteczki 3-fosforanu 1-acyloglicerolu..
Powstały w ten sposób kwas fosfatydowy można hydrolizować przez fosfatazę kwasu fosfatydowego do 1,2-diacyloglicerolu, który można następnie zastosować do syntezy fosfatydylocholiny i fosfatydyloetanoloaminy..
Alternatywny szlak produkcji kwasu fosfatydowego, który obejmuje „recykling” cząsteczek 1,2-diacyloglicerolu, wiąże się z udziałem specyficznych enzymów kinaz, które przenoszą grupy fosforanowe na węgiel w pozycji 3 diacyloglicerolu.
Innym jest hydroliza innych fosfolipidów, katalizowana przez enzymy znane jako fosfolipazy. Przykładem tego procesu jest produkcja kwasu fosfatydowego z fosfatydylocholiny dzięki działaniu fosfolipazy D, która hydrolizuje wiązanie między choliną a grupą fosforanową 3-fosforanu 1,2-diacyloglicerolu..
Produkcja kwasu fosfatydowego w roślinach jest związana z czterema różnymi przedziałami komórek roślinnych: plastydem, retikulum endoplazmatycznym, mitochondriami i kompleksem Golgiego..
Pierwszy etap trasy jest taki sam, jak opisano poprzednio, a w każdym przedziale bierze udział acylotransferaza glicerolu 3-fosforanu, aby przenieść aktywowaną grupę acylo-CoA na węgiel 1 cząsteczki 3-fosforanu glicerolu..
Syntezę kończy enzym zwany acylotransferazą kwasu lizofosfatydowego po przeniesieniu innej grupy acylowej na pozycję C3 kwasu lizofosfatydowego..
W plastydach roślin enzym ten selektywnie przenosi nasycone kwasy tłuszczowe o długości odpowiadającej 16 atomom węgla. Jest to szczególna cecha lipidów syntetyzowanych w tych organellach..
Kwas fosfatydowy jest prekursorem fosfolipidów dla wielu fosfolipidów, galaktolipidów i trójglicerydów w wielu organizmach. Dlatego jest niezbędną cząsteczką dla komórek, mimo że nie spełnia bezpośrednich funkcji strukturalnych..
U zwierząt jeden z produktów jego hydrolizy enzymatycznej, 1,2-diacyloglicerol, jest używany do tworzenia triacyloglicerydów lub triglicerydów poprzez transestryfikację trzecią aktywowaną cząsteczką kwasu tłuszczowego (związaną z częścią CoA).
Triglicerydy są ważnymi cząsteczkami rezerw energii dla zwierząt, ponieważ utlenianie obecnych w nich kwasów tłuszczowych prowadzi do uwolnienia dużych ilości energii oraz prekursorów i produktów pośrednich w innych szlakach metabolicznych..
Inny produkt jego hydrolizy, kwas lizofosfatydowy, jest ważnym drugim przekaźnikiem w niektórych pozakomórkowych szlakach sygnałowych, które obejmują wiązanie się z receptorami na powierzchni innych komórek, między innymi zaangażowanych w powstawanie guzów, angiogenezę i odpowiedzi immunologiczne..
Jego funkcje jako cząsteczki sygnalizacyjnej obejmują udział w indukcji proliferacji komórek, zmniejszonej apoptozy, agregacji płytek krwi, skurczu mięśni gładkich, chemotaksji, inwazji komórek nowotworowych i innych..
U bakterii kwas fosfatydowy jest niezbędny podczas procesu wymiany fosfolipidów na błonie, który dostarcza komórce „osmoprotekcyjne” cząsteczki zwane „oligosacharydami pochodzącymi z błon”..



Jeszcze bez komentarzy