Plik Enzymy restrykcyjne są endonukleazami używanymi przez niektóre archeony i bakterie do zahamowania lub „ograniczenia” rozprzestrzeniania się w nich wirusów. Są szczególnie powszechne w bakteriach i są częścią ich systemu obronnego przed obcym DNA, znanym jako system restrykcji / modyfikacji..
Enzymy te katalizują rozszczepianie dwupasmowego DNA w określonych miejscach, w sposób powtarzalny i bez użycia dodatkowej energii. Większość wymaga obecności kofaktorów, takich jak magnez lub inne dwuwartościowe kationy, chociaż niektóre wymagają również ATP lub S-adenozylometioniny.
Endonukleazy restrykcyjne zostały odkryte w 1978 roku przez Daniela Nathansa, Arbera Wernera i Hamiltona Smitha, którzy za ich odkrycie otrzymali Nagrodę Nobla w dziedzinie medycyny. Ich nazwa generalnie pochodzi od organizmu, na którym zostały po raz pierwszy zaobserwowane.
Takie enzymy są szeroko stosowane w opracowywaniu metod klonowania DNA i innych strategii biologii molekularnej i inżynierii genetycznej. Ich specyficzne cechy rozpoznawania sekwencji i możliwość cięcia sekwencji w pobliżu miejsc rozpoznawania sprawiają, że są one potężnymi narzędziami do eksperymentów genetycznych..
Fragmenty wygenerowane przez enzymy restrykcyjne, które oddziałują na określoną cząsteczkę DNA, można wykorzystać do odtworzenia „mapy” oryginalnej cząsteczki, wykorzystując informacje o miejscach, w których enzym tnie DNA..
Niektóre enzymy restrykcyjne mogą mieć to samo miejsce rozpoznawane w DNA, ale niekoniecznie przecinają je w ten sam sposób. Tak więc istnieją enzymy, które tną pozostawiając tępe końce i enzymy, które tną pozostawiając spójne końce, które mają różne zastosowania w biologii molekularnej.
Obecnie na rynku dostępne są setki różnych enzymów restrykcyjnych oferowanych przez różne domy handlowe; enzymy te pełnią funkcję „niestandardowych” molekularnych nożyczek do różnych celów.
Indeks artykułów
Enzymy restrykcyjne pełnią odwrotną funkcję polimerazy, ponieważ hydrolizują lub przerywają wiązanie estrowe w obrębie wiązania fosfodiestrowego między sąsiednimi nukleotydami w łańcuchu nukleotydowym..
W biologii molekularnej i inżynierii genetycznej są szeroko stosowanymi narzędziami do budowy wektorów ekspresyjnych i do klonowania, a także do identyfikacji określonych sekwencji. Są również przydatne do budowy rekombinowanych genomów i mają duży potencjał biotechnologiczny..
Ostatnie postępy w terapii genowej sprawiają, że obecnie enzymy restrykcyjne są wprowadzane do wektorów, które są nośnikami transportu takich genów do żywych komórek i prawdopodobnie mają zdolność wstawiania do genomu komórkowego w celu dokonania trwałych zmian..
Enzymy restrykcyjne mogą katalizować dwupasmowe cięcie DNA, chociaż niektóre są zdolne do rozpoznawania jednopasmowych sekwencji DNA, a nawet RNA. Cięcie następuje po rozpoznaniu sekwencji.
Mechanizm działania polega na hydrolizie wiązania fosfodiestrowego pomiędzy grupą fosforanową a dezoksyrybozą w szkielecie każdej nici DNA. Wiele enzymów jest zdolnych do cięcia w tym samym miejscu, które rozpoznają, podczas gdy inne tną od 5 do 9 par zasad przed lub po tym samym.
Zwykle te enzymy przecinają na końcu 5 'grupy fosforanowej, dając początek fragmentom DNA z końcem 5' fosforylowym i końcem hydroksylowym 3 '..
Ponieważ białka nie wchodzą w bezpośredni kontakt z miejscem rozpoznawanym w DNA, muszą być przemieszczane sukcesywnie, aż do osiągnięcia określonego miejsca, być może za pomocą mechanizmów „ślizgania” na nici DNA..
Podczas cięcia enzymatycznego wiązanie fosfodiestrowe każdej z nici DNA znajduje się w jednym z aktywnych miejsc enzymów restrykcyjnych. Kiedy enzym opuszcza miejsce rozpoznawania i rozszczepiania, czyni to poprzez nieswoiste przejściowe skojarzenia.
Obecnie znanych jest pięć typów enzymów restrykcyjnych. Oto krótki opis każdego z nich:
Te enzymy to duże pentameryczne białka z trzema podjednostkami, jedną do restrykcji, jedną do metylacji i jedną do rozpoznawania sekwencji w DNA. Te endonukleazy są wielofunkcyjnymi białkami zdolnymi do katalizowania reakcji restrykcji i modyfikacji, mają aktywność ATPazy, a także topoizomerazę DNA..
Enzymy tego typu były pierwszymi odkrytymi endonukleazami, zostały po raz pierwszy oczyszczone w latach 60. XX wieku i od tamtej pory były dogłębnie badane..
Enzymy typu I nie są szeroko stosowane jako narzędzie biotechnologiczne, ponieważ miejsce rozszczepienia może znajdować się w zmiennej odległości do 1000 par zasad od miejsca rozpoznania, co czyni je niewiarygodnymi pod względem odtwarzalności eksperymentalnej..
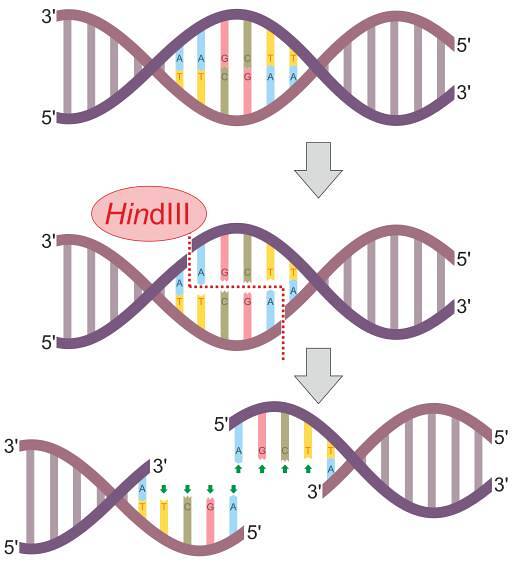
Są to enzymy złożone z homodimerów lub tetramerów, które tną DNA w określonych miejscach o długości od 4 do 8 pz. Te miejsca rozszczepienia są zazwyczaj palindromiczne, to znaczy rozpoznają sekwencje odczytywane w ten sam sposób w obu kierunkach..
Wiele enzymów restrykcyjnych typu II w bakteriach tnie DNA, gdy rozpoznają jego obcy charakter, ponieważ nie ma on typowych modyfikacji, jakie powinno mieć jego własne DNA.
Są to najprostsze enzymy restrykcyjne, ponieważ nie wymagają żadnego innego kofaktora niż magnez (Mg +) do rozpoznawania i cięcia sekwencji DNA..
Precyzja enzymów restrykcyjnych typu II w rozpoznawaniu i wycinaniu prostych sekwencji w DNA w precyzyjnych pozycjach sprawia, że są one jednymi z najpowszechniej stosowanych i nieodzownych w większości dziedzin biologii molekularnej..
W grupie enzymów restrykcyjnych typu II istnieje wiele podklas sklasyfikowanych według pewnych właściwości, które są unikalne dla każdej z nich. Klasyfikacji tych enzymów dokonuje się poprzez dodanie liter alfabetu od A do Z po nazwie enzymu.
Niektóre z podklas najbardziej znanych ze swojej przydatności to:
Są dimerami różnych podjednostek. Rozpoznają sekwencje asymetryczne i są wykorzystywane jako idealne prekursory do wytwarzania enzymów tnących.
Składają się z jednego lub więcej dimerów i pociętego DNA po obu stronach rozpoznawanej sekwencji. Wytnij obie nici DNA w odstępach par zasad przed miejscem rozpoznania.
Enzymy tego typu to polipeptydy z funkcjami podziału i modyfikacji nici DNA. Enzymy te przecinają asymetrycznie obie nici.
Enzymy z tej podklasy są najczęściej używane w inżynierii genetycznej. Mają miejsce katalityczne i generalnie wymagają allosterycznego efektora. Enzymy te muszą współdziałać z dwiema kopiami ich sekwencji rozpoznawania, aby przeprowadzić wydajne cięcie. W tej podklasie znajdują się enzymy EcoRII i EcoRI.
Endonukleazy restrykcyjne typu III składają się tylko z dwóch podjednostek, z których jedna jest odpowiedzialna za rozpoznawanie i modyfikację DNA, a druga za rozszczepianie sekwencji.
Enzymy te do swoich funkcji wymagają dwóch kofaktorów: ATP i magnezu. Enzymy restrykcyjne tego typu posiadają dwa asymetryczne miejsca rozpoznawania, dokonują translokacji DNA w sposób zależny od ATP i tną go od 20 do 30 pz w sąsiedztwie miejsca rozpoznawania..
Enzymy typu IV są łatwe do zidentyfikowania, ponieważ tną DNA ze śladami metylacji, składają się z kilku różnych podjednostek odpowiedzialnych za rozpoznawanie i cięcie sekwencji DNA. Enzymy te wykorzystują GTP i dwuwartościowy magnez jako kofaktory.
Specyficzne miejsca cięcia obejmują nici nukleotydowe z metylowanymi lub hydroksymetylowanymi resztami cytozyny na jednej lub obu niciach kwasów nukleinowych..
Ta klasyfikacja grupuje razem enzymy typu CRISPER-Cas, które identyfikują i wycinają określone sekwencje DNA z atakujących organizmów. Enzymy Cas używają zsyntetyzowanej wiodącej nici RNA firmy CRISPER do rozpoznawania i atakowania atakujących organizmów.
Enzymy zaliczane do typu V to polipeptydy zbudowane według enzymów typu I, II i II. Potrafią wycinać fragmenty DNA prawie każdego organizmu i mieć szeroki zakres długości. Ich elastyczność i łatwość użycia sprawiają, że enzymy te są obecnie jednym z najczęściej używanych narzędzi w inżynierii genetycznej, obok enzymów typu II..
Enzymy restrykcyjne były używane do wykrywania polimorfizmów DNA, zwłaszcza w badaniach genetyki populacji i badaniach ewolucyjnych z wykorzystaniem mitochondrialnego DNA, w celu uzyskania informacji o szybkości podstawiania nukleotydów..
Obecnie wektory używane do transformacji bakterii do różnych celów mają miejsca wieloklonowania, w których znajdują się miejsca rozpoznawane przez wiele enzymów restrykcyjnych..
Wśród tych enzymów najpopularniejsze są EcoRI, II, III, IV i V, otrzymane i opisane po raz pierwszy z E coli; HindIII z H. influenzae i BamHI z B. amyloliquefaciens.


Jeszcze bez komentarzy