Plik prawo Hardy-Weinberg, Nazywana również zasadą Hardy'ego-Weinberga lub równowagą, składa się z twierdzenia matematycznego opisującego hipotetyczną populację diploidalną z rozmnażaniem płciowym, która nie ewoluuje - częstotliwości alleli nie zmieniają się z pokolenia na pokolenie.
Zasada ta zakłada pięć warunków niezbędnych do utrzymania populacji na stałym poziomie: brak przepływu genów, brak mutacji, losowe kojarzenie, brak doboru naturalnego i nieskończenie duży rozmiar populacji. W ten sposób przy braku tych sił populacja pozostaje w równowadze.
Gdy którekolwiek z powyższych założeń nie zostanie spełnione, następuje zmiana. Z tego powodu dobór naturalny, mutacje, migracje i dryf genetyczny to cztery mechanizmy ewolucyjne..
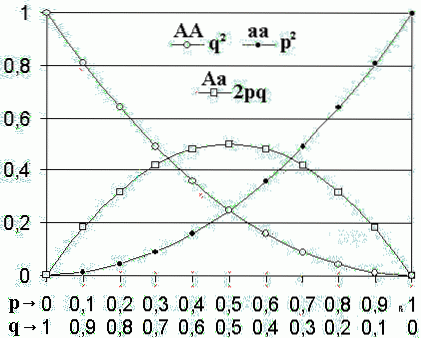
Zgodnie z tym modelem, gdy częstości alleli populacji są p Y co, częstotliwości genotypów będą pdwa, dwapq Y codwa.
Możemy zastosować równowagę Hardy'ego-Weinberga do obliczania częstotliwości pewnych interesujących nas alleli, na przykład w celu oszacowania odsetka heterozygot w populacji ludzkiej. Możemy również zweryfikować, czy populacja jest w równowadze, czy też nie, i zaproponować hipotezy, że siły działają na tę populację..
Indeks artykułów
Zasada Hardy'ego-Weinberga narodziła się w 1908 roku i swoją nazwę zawdzięcza swoim naukowcom G.H. Hardy i W. Weinberg, którzy niezależnie doszli do tych samych wniosków.
Wcześniej inny biolog imieniem Udny Yule zajął się tym problemem w 1902 roku. Yule rozpoczął od zestawu genów, w których częstości występowania obu alleli wynosiły 0,5 i 0,5. Biolog wykazał, że częstotliwości utrzymywały się przez kolejne pokolenia.
Chociaż Yule doszedł do wniosku, że częstotliwości alleli można utrzymać na stałym poziomie, jego interpretacja była zbyt dosłowna. Uważał, że jedyny stan równowagi występuje, gdy częstotliwości odpowiadają wartości 0,5.
Yule gorąco dyskutowała o swoich nowych odkryciach z R.C. Punnett - szeroko znany w dziedzinie genetyki z wynalezienia słynnego „placu Punnetta”. Chociaż Punnett wiedział, że Yule się mylił, nie znalazł matematycznego sposobu, aby to udowodnić..
Dlatego Punnett skontaktował się ze swoim przyjacielem matematykiem Hardym, któremu udało się rozwiązać go natychmiast, powtarzając obliczenia przy użyciu zmiennych ogólnych, a nie stałej wartości 0,5, jak to zrobił Yule..
Genetyka populacji ma na celu zbadanie sił, które prowadzą do zmian częstotliwości alleli w populacjach, integrując teorię ewolucji Karola Darwina przez dobór naturalny i genetykę mendlowską. Dziś jej zasady stanowią podstawę teoretyczną do zrozumienia wielu aspektów biologii ewolucyjnej..
Jedną z kluczowych idei genetyki populacyjnej jest związek między zmianami we względnej obfitości cech a zmianami we względnej obfitości alleli, które ją regulują, co wyjaśnia zasada Hardy'ego-Weinberga. W rzeczywistości to twierdzenie zapewnia ramy koncepcyjne genetyki populacyjnej..
W świetle genetyki populacji koncepcja ewolucji jest następująca: zmiana częstości alleli na przestrzeni pokoleń. Gdy nie ma zmiany, nie ma ewolucji.
Równowaga Hardy'ego-Weinberga to model zerowy, który pozwala nam określić zachowanie częstotliwości genów i alleli w różnych pokoleniach. Innymi słowy, jest to model opisujący zachowanie genów w populacjach w szeregu określonych warunków..
W twierdzeniu Hardy'ego-Weinbergma częstość alleliczna DO (allel dominujący) jest reprezentowany przez literę p, podczas gdy częstotliwość alleli do (allel recesywny) jest reprezentowany przez literę co.
Oczekiwane częstości genotypów to pdwa, dwapq Y codwa, dla homozygotycznej dominanty (AA), heterozygotyczny (Aa) i homozygotyczny recesywny (aa), odpowiednio.
Jeśli w tym miejscu są tylko dwa allele, suma częstotliwości tych dwóch alleli musi koniecznie wynosić 1 (p + q = 1). Rozwinięcie dwumianowe (p + q)dwa reprezentują częstotliwości genotypów pdwa + dwapq + qdwa = 1.
W populacji składające się na nią osobniki krzyżują się, aby dać potomstwo. Ogólnie rzecz biorąc, możemy wskazać najważniejsze aspekty tego cyklu reprodukcyjnego: produkcję gamet, ich fuzję, aby dać początek zygocie oraz rozwój zarodka, aby dać początek nowej generacji..
Wyobraź sobie, że możemy prześledzić mendlowski proces genowy we wspomnianych wydarzeniach. Robimy to, ponieważ chcemy wiedzieć, czy i dlaczego allel lub genotyp zwiększy lub zmniejszy częstotliwość..
Aby zrozumieć, w jaki sposób częstotliwości genów i alleli różnią się w populacji, prześledzimy produkcję gamet przez zestaw myszy. W naszym hipotetycznym przykładzie krycie następuje losowo, w którym wszystkie plemniki i komórki jajowe są losowo mieszane..
W przypadku myszy to założenie nie jest prawdziwe i jest jedynie uproszczeniem ułatwiającym obliczenia. Jednak w niektórych grupach zwierząt, takich jak niektóre szkarłupnie i inne organizmy wodne, gamety są wydalane i zderzają się losowo..
Skoncentrujmy teraz naszą uwagę na konkretnym locus z dwoma allelami: DO Y do. Zgodnie z prawem ogłoszonym przez Gregora Mendla, każda gameta otrzymuje allel z locus A. Załóżmy, że 60% zalążków i plemników otrzymuje allel DO, podczas gdy pozostałe 40% otrzymało allel do.
Stąd częstotliwość alleli DO wynosi 0,6 i allelu do wynosi 0,4. Ta grupa gamet zostanie przypadkowo znaleziona, aby dać początek zygocie. Jakie jest prawdopodobieństwo, że utworzą one każdy z trzech możliwych genotypów? Aby to zrobić, musimy pomnożyć prawdopodobieństwa w następujący sposób:
Genotyp AA: 0,6 x 0,6 = 0,36.
Genotyp Aa: 0,6 x 0,4 = 0,24. W przypadku heterozygoty istnieją dwie formy, w których może powstać. Pierwszy, w którym plemnik niesie allel DO a zalążek allelu do, lub odwrotnie, plemniki do i komórka jajowa DO. Dlatego dodajemy 0,24 + 0,24 = 0,48.
Genotyp aa: 0,4 x 0,4 = 0,16.
Teraz wyobraźmy sobie, że te zygoty rozwijają się i stają się dorosłymi myszami, które ponownie będą wytwarzać gamety, czy oczekiwalibyśmy, że częstotliwości alleli będą takie same lub różne od poprzedniej generacji?
Genotyp AA wyprodukuje 36% gamet, podczas gdy heterozygoty wyprodukują 48% gamet i genotyp aa 16%.
Aby obliczyć częstotliwość nowego allelu, dodajemy częstotliwość homozygot plus połowa heterozygot, w następujący sposób:
Częstotliwość alleli DO: 0,36 + ½ (0,48) = 0,6.
Częstotliwość alleli do: 0,16 + ½ (0,48) = 0,4.
Jeśli porównamy je z częstotliwościami początkowymi, zdamy sobie sprawę, że są identyczne. Dlatego zgodnie z koncepcją ewolucji, ponieważ nie ma zmian w częstości alleli na przestrzeni pokoleń, populacja jest w równowadze - nie ewoluuje..
Jakie warunki musi spełnić poprzednia populacja, aby częstości jej alleli pozostały niezmienne przez pokolenia? W modelu równowagi Hardy'ego-Weinberga populacja, która nie ewoluuje, spełnia następujące założenia:
Populacja musi być bardzo duża, aby uniknąć stochastycznych lub losowych skutków dryfu genów.
Gdy populacje są małe, efekt dryfu genów (losowe zmiany częstości alleli, z jednego pokolenia na drugie) z powodu błędu próbkowania jest znacznie większy i może prowadzić do utrwalenia lub utraty niektórych alleli..
Migracje nie występują w populacji, więc allele, które mogą zmieniać częstość genów, nie mogą przybywać ani opuszczać.
Mutacje to zmiany w sekwencji DNA i mogą mieć różne przyczyny. Te losowe zmiany modyfikują pulę genów w populacji, wprowadzając lub eliminując geny w chromosomach.
Mieszanie gamet musi odbywać się losowo - tak jak w założeniu, którego użyliśmy w przykładzie myszy. Dlatego nie powinno się dokonywać wyboru partnera wśród osobników w populacji, w tym chowu wsobnego (reprodukcja osobników spokrewnionych).
Kiedy kojarzenie nie jest przypadkowe, nie powoduje zmiany częstości alleli z jednego pokolenia na drugie, ale może generować odchylenia od spodziewanych częstości genotypów..
Nie ma zróżnicowanego sukcesu reprodukcyjnego osobników z różnymi genotypami, które mogłyby zmienić częstość alleli w populacji..
Innymi słowy, w hipotetycznej populacji wszystkie genotypy mają takie samo prawdopodobieństwo reprodukcji i przeżycia..
Kiedy populacja nie spełnia tych pięciu warunków, wynikiem jest ewolucja. Logicznie rzecz biorąc, naturalne populacje nie spełniają tych założeń. Dlatego model Hardy'ego-Weinberga jest używany jako hipoteza zerowa, która pozwala nam dokonać przybliżonych oszacowań genu i częstotliwości allelicznych..
Oprócz braku tych pięciu warunków istnieją inne możliwe przyczyny, dla których populacja nie jest w równowadze.
Jeden z nich występuje, gdy loci są związane z płcią lub zjawiskiem zniekształcenia w segregacji lub napęd mejotyczny (gdy każda kopia genu lub chromosomu nie jest przekazywana z równym prawdopodobieństwem następnemu pokoleniu).
Szacuje się, że w Stanach Zjednoczonych jeden na 10 000 noworodków cierpi na chorobę zwaną fenyloketonurią..
Zaburzenie to występuje tylko u homozygot recesywnych w zaburzeniu metabolicznym. Znając te dane, jaka jest częstotliwość nosicieli choroby w populacji??
Aby zastosować równanie Hardy'ego-Weinberga musimy założyć, że wybór partnera nie jest związany z genem związanym z patologią i nie ma chowu wsobnego.
Ponadto zakładamy, że w Stanach Zjednoczonych nie ma zjawisk migracyjnych, nie ma nowych mutacji w fenyloketonurii, a prawdopodobieństwo reprodukcji i przeżycia jest takie samo dla genotypów..
Jeśli powyższe warunki są spełnione, możemy użyć równania Hardy'ego-Weinberga do wykonania obliczeń związanych z problemem..
Wiemy, że jeden przypadek choroby występuje co 10 000 urodzeń, więc codwa = 0,0001, a częstotliwość allelu recesywnego będzie pierwiastkiem kwadratowym z tej wartości: 0,01.
Co p = 1 - co, musimy p wynosi 0,99. Teraz mamy częstotliwość obu alleli: 0,01 i 0,99. Częstotliwość nośna odnosi się do częstotliwości heterozygot, którą oblicza się jako 2pq. Tak więc 2pq = 2 x 0,99 x 0,01 = 0,0198.
Odpowiada to około 2% populacji. Pamiętaj, że to tylko przybliżony wynik.
Jeśli znamy liczbę każdego genotypu w populacji, możemy wywnioskować, czy znajduje się on w równowadze Hardy'ego-Weinberga. Kroki umożliwiające rozwiązanie tego typu problemów są następujące:
p = D. + ½ H.
q = R + ½ H.
Na przykład chcemy sprawdzić, czy następująca populacja motyli jest w równowadze Hardy'ego-Weinberga: jest 79 osobników o homozygotycznym dominującym genotypie (AA), 138 heterozygoty (Aa) i 61 homozygot recesywnych (aa).
Pierwszym krokiem jest obliczenie obserwowanych częstotliwości. Robimy to, dzieląc liczbę osobników na genotyp przez całkowitą liczbę osobników:
D = 79/278 = 0,28
H = 138/278 = 0,50
R = 61/278 = 0,22
Aby sprawdzić, czy dobrze się spisałem, pierwszym krokiem jest dodanie wszystkich częstotliwości i powinno dać 1.
Drugim krokiem jest obliczenie częstotliwości alleli.
p = 0,28 + ½ (0,50) = 0,53
co = 0,22 + ½ (0,50) = 0,47
Na podstawie tych danych mogę obliczyć spodziewane częstości występowania genotypów (pdwa, dwadlaczego i codwa)
pdwa = 0,28
dwapq = 0,50
codwa = 0.22
Obliczam oczekiwane liczby, mnożąc oczekiwane częstotliwości przez liczbę osobników. W tym przypadku liczba obserwowanych i oczekiwanych osobników jest identyczna, więc mogę stwierdzić, że populacja jest w równowadze.
Gdy otrzymane liczby nie są identyczne, muszę zastosować wspomniany test statystyczny (Xdwa od Pearson).



Jeszcze bez komentarzy