Plik miozyna jest to motor molekularny, z natury białkowy, zdolny do poruszania się po filamentach aktyny w cytozolu. Energia, która napędza wypieranie miozyny, pochodzi z hydrolizy ATP. Z tego powodu miozyna jest często definiowana jako enzym mechanochemiczny.
U eukariotów miozyna jest białkiem występującym w bardzo dużej ilości. Istnieją różne klasy miozyny, które są kodowane przez rodzinę genów. W drożdżach wyróżnia się 5 klas, u ssaków opisano kilkadziesiąt.
Miozyna ma wiele funkcji. Miozyna I wraz z aktyną uczestniczy w ruchu keratocytów.
Miozyna II zapewnia sztywność błony komórkowej, bierze udział w cytokinezy i skurczu mięśni. Obie miozyny I i II współpracują z migracją komórek. Miozyny I i V przenoszą pęcherzyki wzdłuż włókien aktyny.
Indeks artykułów
W mikroskopach elektronowych typowa struktura izoform miozyny ma trzy domeny: głowę, szyję i ogon. W wyniku hydrolizy z chymotrypsyną uzyskuje się segment składający się z głowy i szyi, zwany ciężką meromiozyną (HMM) oraz odcinek ogona, zwany lekką meromiozyną (LMM).
Domena głowy jest N-końcowym końcem łańcucha ciężkiego, a domena ogonowa jest C-końcem łańcucha lekkiego.
Klasy miozyny można rozróżnić na podstawie liczby łańcuchów polipeptydowych, które ją tworzą, oraz obfitości i klasy lekkich łańcuchów przyczepionych do szyi.
Miozyna I ma łańcuch polipeptydowy, który tworzy głowę, a na jej ogonie brakuje regionów alfa-helikalnych. Podczas gdy miozyny I i V mają dwa łańcuchy polipeptydowe, a zatem tworzą dwie głowy i ogon, w którym spiralne łańcuchy alfa są zwinięte, tworząc strukturę podobną do pręta.
Miozyny I i V mają miejsca wiązania kalmoduliny, która reguluje i wiąże Ca+dwa, na łańcuchach lekkich. Myosin I naprawia Ca+dwa w lekkich łańcuchach, ale robi to inaczej niż kalmodulina.
Na poziomie mechanochemicznym miozyny mają trzy cechy, a mianowicie:
- Głowa miozyny jest domeną motoryczną, która rozwija się w dyskretnych krokach: połączenie głowy miozyny z filamentem aktyny, jej nachylenie i późniejsza separacja powodują ruch miozyny. Ten proces jest cykliczny i zależy od ATP.
- Zmiany konformalne: hydroliza cząsteczki ATP jest sprzężona z każdym etapem cząsteczki miozyny poprzez poziomy amplifikacji i transmisji. Wiąże się to z dużymi zmianami konformacyjnymi miozyny..
Pierwszy poziom amplifikacji jest spowodowany utratą grupy gamma-fosforanowej ATP, co umożliwia reorganizację elementów strukturalnych w miejscu wiązania ATP. Ta reorganizacja jest skoordynowana ze zmianami strukturalnymi w miejscu wiązania aktyny..
Drugi poziom amplifikacji obejmuje przekazywanie zmiany konformacyjnej w miejscu aktywnym do składników strukturalnych końca karboksylowego..
- Kierunkowość: Stwierdzono, że miozyny mają polaryzację lub odwrotną kierunkowość w kierunku (+) końca filamentu aktyny. Ten wniosek pochodzi z eksperymentów poślizgu filamentów aktyny przy użyciu fluorescencyjnego mikroskopu świetlnego..
Miozyna wraz z aktyną bierze udział m.in. w skurczach mięśni, adhezji komórek, cytokinezie, usztywnianiu błon korowych i przemieszczaniu niektórych pęcherzyków..
Wady miozyny mogą powodować stany patologiczne. Na przykład defekty miozyn I i V są związane, odpowiednio, z miopatiami miozyny i zaburzeniami pigmentacji (zespół Griscelli). Podczas gdy zaburzenia w izoformach miozyny VI powodują utratę słuchu.
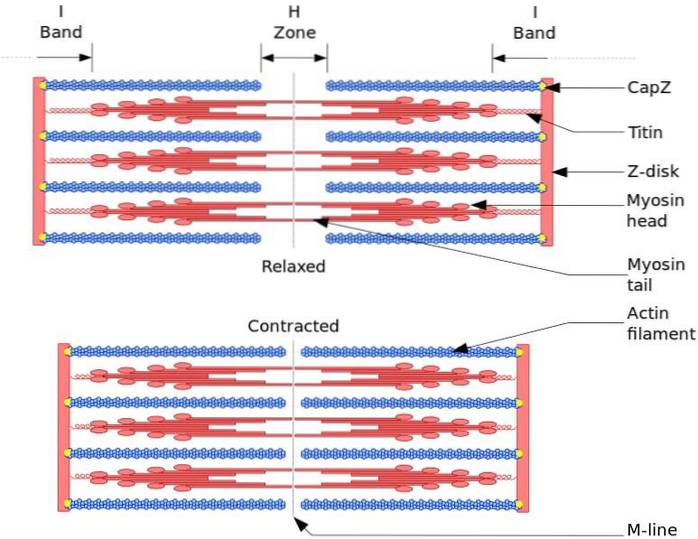
Funkcjonalną i strukturalną jednostką mięśni szkieletowych jest sarkomer. Podczas skurczu mięśni długość sarkomeru sięga 30% jego pierwotnej długości.
Sarcomeres składa się z grubych włókien miozyny i cienkich włókien aktyny, które są zorganizowane w złożony sposób. Ogólnie rzecz biorąc, głowy miozyny znajdują się na dalszych końcach włókna, a ich ogony w kierunku środka sarkomeru, a organizacja jest dwubiegunowa.
Aby wystąpił skurcz mięśni, głowy miozyny na przeciwnych końcach muszą przesunąć się w kierunku dysku Z lub (+) końca włókna. Ponieważ organizacja grubych włókien jest dwubiegunowa, następuje przesuwanie cienkich włókien na grubych włóknach, napędzane przez ATP.
Siła przemieszczenia występuje, ponieważ setki główek miozyny z grubego włókna oddziałują z cienkim włóknem.
Podczas mitozy, gdy mikrotubule na biegunach wrzeciona rozdzielają się, aktyna i miozyna II tworzą kurczliwy pierścień na równiku komórki. Ten pierścień kurczy się, zmniejszając jego średnicę i dzieląc komórkę na dwie części..
W zmutowanych komórkach pozbawionych miozyny II błona plazmatyczna łatwo ulega deformacji po przyłożeniu siły zewnętrznej. Dzieje się tak, ponieważ miozyna II zapewnia siłę agregacji białkom błony komórkowej..
W tkance nabłonkowej kurczliwe wiązki aktyny i miozyny II znajdują się w pobliżu błony komórkowej i tworzą kolistą obwódkę otaczającą wewnętrzną powierzchnię komórki. Ta okrągła opaska określa kształt komórki i utrzymuje więź między komórkami..
Kontakt między komórkami zachodzi poprzez zjednoczenie okrągłego pasa z komórkowymi cząsteczkami adhezyjnymi za pomocą białek unii.
Dowody eksperymentalne pokazują, że miozyna V przenosi błonę z aparatu Golgiego na obrzeże komórki. Oto niektóre dowody:
- W komórkach tkanki nerwowej za pomocą immunofluorescencji astrocytów stwierdzono, że miozyna V znajduje się obok aparatu Golgiego..
- U drożdży mutacje w genie miozyny V zakłócają wydzielanie białka iw konsekwencji białko gromadzi się w cytozolu.
- Izoformy miozyny I są odpowiedzialne za transport wakuoli w kierunku błony komórkowej. Używając specyficznych przeciwciał przeciwko izoformom miozyny I, stwierdzono, że te izoformy są zlokalizowane w różnych częściach komórki..
Na przykład, gdy żywa ameba jest wyznakowana przeciwciałem przeciwko IC miozyny, transport wakuoli do błony zostaje zatrzymany. Z tego powodu wakuola rozszerza się, a komórka pęka.
Istnieje wiele genów i mutacji, które powodują utratę słuchu. Ta choroba jest często monogeniczna .
Niekonwencjonalne mutacje miozyny, z jedną lub dwiema główkami miozyny, wpływają na funkcjonowanie ucha wewnętrznego. Niektóre ze zmutowanych izoform miozyny to miozyna IIIA, miozyna VIIA i miozyna XVA. Niedawno odkryto dwie mutacje w miozynie VI.
Mutacje w miozynie VI to c.897G> T i p.926Q. Pierwsza mutacja wpływa na region, który oddziałuje z miejscem aktywnym, zwany Switch I. Homozygotycy mutacji wcześnie przejawiają fenotyp, powodując poważne skutki.
Druga mutacja wpływa na region naładowanych reszt w alfa helisie w ogonie miozyny VI. Region ten jest ważny dla dimeryzacji proksymalnej motorycznej i wpływa na funkcję stereo-rzęskową miozyny VI..
Inną mutacją jest p.Asn207Ser, która wytwarza silnik niezdolny do wytworzenia siły. Dzieje się tak, ponieważ Asn 207 jest resztą aminokwasową miejsca aktywnego, której funkcją jest wiązanie i hydroliza ATP.
Mutacja p.Arg657Trp powoduje utratę funkcji miozyny VI. Reszta Arg bierze udział w zmianach konformacyjnych, które wiążą hydrolizę z ruchem miozyny.
Myosin X (Myo10) to niekonwencjonalna miozyna, która ulega ekspresji w mózgu, śródbłonku i wielu nabłonkach. Myo10 i trzy klasy projekcji opartych na aktynie (filopodia, invadopodia i filopodia-like projeions) działają podczas przerzutów raka.
Inwazyjne komórki rakowe mają dużą liczbę filopodiów i wyrażają wysoki poziom fascyny. To białko tworzy wiązania poprzeczne między filamentami aktyny. Aby uciec od pierwotnego guza, powstają naciekające obfitujące w aktywność proteolityczną, które trawią otaczającą macierz zewnątrzkomórkową..
Gdy komórki dotrą do macierzy zewnątrzkomórkowej, wypustki podobne do filopodiów pomagają rozproszyć się i skolonizować. Wysokie poziomy Myo10 wskazują na wysoką agresywność i przerzuty w raku piersi.
Wyciszenie MyoX powoduje utratę przerzutowego charakteru komórek, które nie są w stanie tworzyć wypustek opartych na aktynie. Wszystkie te wypustki mają zrosty oparte na integrynie, które są przenoszone przez Myo10 w filopodium..
MyoX bierze udział w tworzeniu centrosomu. Brak MyoX sprzyja tworzeniu się wrzecion wielobiegunowych. MyoX bierze również udział w sygnalizacji w komórkach nowotworowych. Na przykład MyoX jest aktywowany przez trifosforan 3,4,5-inozytolu (PIP3).



Jeszcze bez komentarzy