
Plik teoria fałdowania membrany proponuje, że błony organelli pochodzą z ekspansji i wgłębienia błony komórkowej. J.D Robertson, pionier mikroskopii elektronowej, zauważył w 1962 roku, że kilka ciał wewnątrzkomórkowych posiadało strukturę wizualnie identyczną z błoną plazmatyczną..
Idea struktury, która rozgranicza komórki, pojawiła się natychmiast po pojawieniu się koncepcji „komórki”, dla której przeprowadzono kilka badań w celu wyjaśnienia właściwości tej struktury..
Indeks artykułów
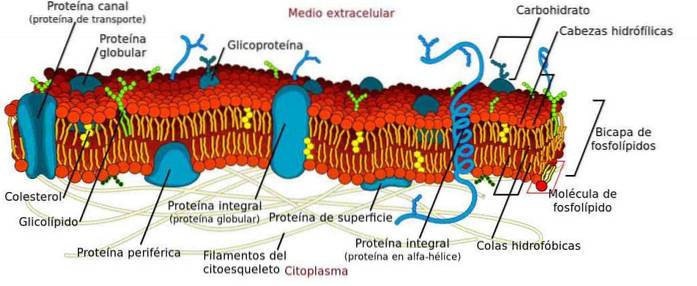
Błona plazmatyczna jest strukturą utworzoną przez podwójną warstwę fosfolipidów zorganizowanych w taki sposób, że grupy polarne są zorientowane w kierunku cytozolu i ośrodka zewnątrzkomórkowego, podczas gdy grupy niepolarne są zorganizowane w kierunku wnętrza błony..
Jego główną funkcją jest definiowanie komórek, zarówno eukariotycznych, jak i prokariotycznych, ponieważ fizycznie oddziela cytoplazmę od środowiska zewnątrzkomórkowego..
Pomimo swojej funkcji strukturalnej dobrze wiadomo, że membrana nie jest statyczna, ale raczej elastyczną i dynamiczną barierą, w której zachodzi wiele procesów niezbędnych dla komórki..
Niektóre procesy zachodzące w błonie to zakotwiczenie cytoszkieletu, transport cząsteczek, przekazywanie sygnałów i łączenie się z innymi komórkami w celu wytworzenia tkanek. Ponadto wiele organelli ma również błonę, w której zachodzą inne procesy o wielkim znaczeniu..
Na długo przed tym, jak Robertson zaproponował teorię fałdowania membrany w 1962 r., Po raz pierwszy przeprowadzono badania, aby określić, jak wygląda ta struktura. W przypadku braku mikroskopu elektronowego przeważały badania elektrofizjologiczne, wśród których znalazły się:
Overton zauważył, że lipidy łatwiej przenikały przez błonę komórkową niż cząsteczki innego rodzaju, dlatego wywnioskował, że błona musi składać się w większości z lipidów..
J. Bernstein przedstawił swoją hipotezę, w której wspomniał, że komórki składają się z roztworu z wolnymi jonami ograniczonymi cienką warstwą nieprzepuszczalną dla wspomnianych naładowanych cząsteczek..
Fricke zmierzył zdolność błony erytrocytów do przechowywania ładunków (pojemność), określając, że ta wartość wynosiła 0,81 µF / cmdwa.
Później ustalono, że membrany innych typów komórek mają podobne wartości pojemności, dlatego membrana powinna być strukturą jednolitą.
Gorter i Grendel zmierzyli powierzchnię erytrocytów ssaków za pomocą mikroskopu. Następnie wyekstrahowali lipidy ze znanej liczby komórek tego typu i zmierzyli zajmowaną przez nie powierzchnię..
W rezultacie uzyskali stosunek komórki do błony wynoszący 1: 2. Oznaczało to, że błona komórkowa była podwójną strukturą, co dało początek terminowi „dwuwarstwowy lipid”.
Badania przeprowadzone przed rokiem 1935 sugerowały obecność białek w błonie, co skłoniło Danielli i Davson do zaproponowania modelu sandwich lub modelu białko-lipid-białko..
Zgodnie z tym modelem błona plazmatyczna składa się z dwóch warstw fosfolipidów znajdujących się między dwiema warstwami białek, które są związane z błoną poprzez oddziaływania elektrostatyczne..
W 1959 roku, dzięki pojawieniu się mikroskopii elektronowej, J. David Robertson zebrał wystarczające dowody, aby potwierdzić i uzupełnić modele zaproponowane przez Gortera i Grendela (1925) oraz Danielli i Davson (1935) i zaproponować model "Unitary Membrane"..
Model ten zachowuje charakterystykę modelu dwuwarstwy lipidowej zaproponowanej przez Danielli i Davson, ze zmiennością warstwy białkowej, która w tym przypadku jest asymetryczna i nieciągła. .
Pojawienie się mikroskopii elektronowej pozwoliło mieć dość jasne wyobrażenie o tym, jak powstała błona plazmatyczna.
Jednak temu faktowi towarzyszyła wizualizacja wielu błon wewnątrzcytoplazmatycznych, które tworzyły przedziały wewnątrzkomórkowe, co skłoniło Robertsona w 1962 r. Do zaproponowania „teorii fałdowania błony”..
Teoria fałdowania błony polega na tym, że błona plazmatyczna zwiększyła swoją powierzchnię i została wklęsła, dając początek błonom wewnątrzcytoplazmatycznym, błony te otaczały cząsteczki znajdujące się w cytozolu, tworząc w ten sposób organelle.
Zgodnie z tą teorią, otoczka jądrowa, retikulum endoplazmatyczne, aparat Golgiego, lizosomy i wakuole mogły powstać w ten sposób..
Ciągłość, która istnieje między błoną plazmatyczną a pierwszymi trzema wspomnianymi powyżej organellami, została potwierdzona w badaniach mikroskopii elektronowej w różnych typach komórek..
Jednak Robertson również zaproponował w swojej teorii, że organelle pęcherzykowe, takie jak lizosomy i wakuole, również powstały z wgłębień, które następnie oddzieliły się od błony..
Ze względu na charakterystykę teorii fałdowania membrany uważa się ją za rozszerzenie modelu membrany jednostkowej, który sam zaproponował w 1959 r..
Mikrografie wykonane przez Robertsona pokazują, że wszystkie te membrany są takie same i dlatego powinny mieć dość podobny skład..
Jednak specjalizacja organelli znacznie modyfikuje skład błon, co zmniejsza ich wspólne właściwości na poziomie biochemicznym i molekularnym..
W ten sam sposób, fakt, że membrany pełnią swoją główną funkcję, to służenie jako stabilna bariera dla ośrodków wodnych..
Dzięki wszystkim badaniom przeprowadzonym w latach 1895-1965, a zwłaszcza badaniom mikroskopowym przeprowadzonym przez J.D. Robertson, podkreślono znaczenie błon komórkowych.
Od jej jednolitego modelu zaczęto podkreślać zasadniczą rolę, jaką błona odgrywa w strukturze i funkcji komórek, do tego stopnia, że badanie tej struktury uważa się za fundamentalne zagadnienie we współczesnej biologii..
Jednak w odniesieniu do wkładu teorii fałdowania membrany nie jest to obecnie akceptowane. Jednak w tamtym czasie doprowadziło to do większej liczby ekspertów w tej dziedzinie, którzy próbowali wyjaśnić pochodzenie nie tylko błon komórkowych, ale także pochodzenia samej komórki eukariotycznej, jak uczynił to Lynn Margulis w 1967 r., Podnosząc teorię endosymbiotyczną..



Jeszcze bez komentarzy