Plik fosforylacja oksydacyjna to proces, w którym cząsteczki ATP są syntetyzowane z ADP i P.ja (nieorganiczny fosforan). Mechanizm ten jest wykonywany przez bakterie i komórki eukariotyczne. W komórkach eukariotycznych fosforylacja zachodzi w macierzy mitochondrialnej komórek niefotosyntetycznych.
Produkcja ATP jest napędzana przez transfer elektronów z koenzymów NADH lub FADHdwa do Odwa. Proces ten reprezentuje najwyższą produkcję energii w komórce i pochodzi z rozkładu węglowodanów i tłuszczów..
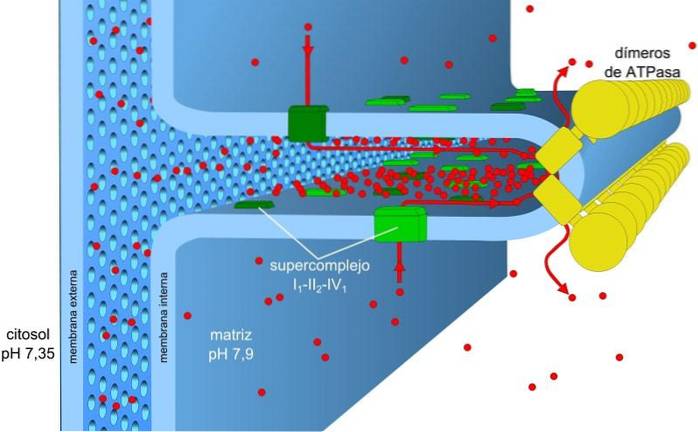
Energia zmagazynowana w gradiencie ładunku i pH, znana również jako siła napędowa protonów, umożliwia ten proces. Generowany gradient protonów powoduje, że zewnętrzna część membrany ma ładunek dodatni ze względu na stężenie protonów (H+), a macierz mitochondrialna jest ujemna.
Indeks artykułów
Z błoną związane są procesy transportu elektronów i fosforylacji oksydacyjnej. U prokariotów mechanizmy te zachodzą przez błonę plazmatyczną. W komórkach eukariotycznych łączą się z błoną mitochondrialną.
Liczba mitochondriów znajdujących się w komórkach różni się w zależności od rodzaju komórki. Na przykład u ssaków erytrocyty nie mają tych organelli, podczas gdy inne typy komórek, takie jak komórki mięśniowe, mogą ich mieć nawet miliony..
Błona mitochondrialna składa się z prostej błony zewnętrznej, nieco bardziej złożonej błony wewnętrznej, a pomiędzy nimi przestrzeni międzybłonowej, w której znajduje się wiele enzymów zależnych od ATP.
Zewnętrzna błona zawiera białko zwane porinem, które tworzy kanały dla prostej dyfuzji małych cząsteczek. Ta membrana jest odpowiedzialna za utrzymanie struktury i kształtu mitochondriów.
Wewnętrzna błona ma większą gęstość i jest bogata w białka. Jest również nieprzepuszczalny dla cząsteczek i jonów, więc aby go przekroczyć, potrzebują białek międzybłonowych do ich transportu..
Wewnątrz matrycy rozciągają się fałdy wewnętrznej membrany, tworząc grzbiety, które pozwalają jej mieć dużą powierzchnię w małej objętości.
Uważa się, że mitochondrium jest głównym producentem energii komórkowej. Zawiera enzymy zaangażowane w procesy cyklu kwasu cytrynowego, utleniania kwasów tłuszczowych oraz enzymy redoks i białka transportu elektronów i fosforylacji ADP.
Gradient stężenia protonów (gradient pH) i gradient ładunku lub potencjał elektryczny w wewnętrznej błonie mitochondriów są odpowiedzialne za siłę napędową protonów. Niska przepuszczalność wewnętrznej membrany dla jonów (innych niż H.+) pozwala mitochondriom mieć stabilny gradient napięcia.
Transport elektroniczny, pompowanie protonów i pozyskiwanie ATP zachodzą jednocześnie w mitochondriach dzięki sile napędowej protonów. Gradient pH utrzymuje kwaśne warunki w międzybłonowej i macierzy mitochondrialnej w warunkach zasadowych.
Na każde dwa elektrony przeniesione do Odwa około 10 protonów jest pompowanych przez membranę, tworząc gradient elektrochemiczny. Energia uwalniana w tym procesie jest wytwarzana stopniowo w wyniku przejścia elektronów przez łańcuch transportowy.
Energia uwalniana podczas reakcji utleniania i redukcji NADH i FADHdwa jest znacznie wysoki (około 53 kcal / mol na każdą parę elektronów), więc aby mógł być wykorzystany do produkcji cząsteczek ATP, musi być wytwarzany stopniowo wraz z przejściem elektronów przez transportery.
Są one zorganizowane w cztery kompleksy zlokalizowane na wewnętrznej błonie mitochondrialnej. Sprzężenie tych reakcji z syntezą ATP odbywa się w piątym kompleksie.
NADH przenosi parę elektronów, które wchodzą do kompleksu I łańcucha transportu elektronów. Elektrony są przenoszone do mononukleotydu flawiny, a następnie do ubichinonu (koenzym Q) przez transporter żelazo-siarka. Ten proces uwalnia dużą ilość energii (16,6 kcal / mol).
Ubichinon przenosi elektrony przez błonę do kompleksu III. W tym kompleksie elektrony przechodzą przez cytochromy bi c1 dzięki transporterowi żelazo-siarka.
Elektrony przechodzą z kompleksu III do kompleksu IV (oksydaza cytochromu c), przenoszone jeden po drugim w cytochromie c (białko błony obwodowej). W kompleksie IV elektrony przechodzą przez parę jonów miedzi (Cudodwa+), a następnie cytochrom cdo, następnie do innej pary jonów miedzi (Cubdwa+) i od tego do cytochromu a3.
Wreszcie elektrony są przenoszone do Odwa który jest ostatnim akceptorem i tworzy cząsteczkę wody (H.dwaO) dla każdej otrzymanej pary elektronów. Przejście elektronów od kompleksu IV do Odwa Generuje również dużą ilość darmowej energii (25,8 kcal / mol).
Kompleks II (reduktaza bursztynianowa CoQ) otrzymuje parę elektronów z cyklu kwasu cytrynowego przez utlenianie cząsteczki bursztynianu do fumaranu. Te elektrony są przenoszone do FAD, a następnie przechodzą przez grupę żelazo-siarkową do ubichinonu. Z tego koenzymu przechodzą do kompleksu III i podążają opisaną wcześniej trasą.
Energia uwolniona w reakcji przeniesienia elektronów do FAD nie wystarcza do przepuszczenia protonów przez błonę, więc na tym etapie łańcucha nie jest generowana żadna siła napędowa protonów, w konsekwencji FADH daje mniej H+ że NADH.
Energia wytworzona w opisanym wcześniej procesie transportu elektronów musi być zdolna do wykorzystania do produkcji ATP, reakcji katalizowanej przez enzym syntazę ATP lub kompleks V.Zachowanie tej energii jest znane jako sprzężenie energii, a mechanizm został opracowany. trudne do scharakteryzowania.
Opisano kilka hipotez opisujących tę transdukcję energii. Najlepiej akceptowaną jest hipoteza sprzężenia chemiosmotycznego, opisana poniżej..
Mechanizm ten sugeruje, że energia wykorzystywana do syntezy ATP pochodzi z gradientu protonów w błonach komórkowych. Proces ten interweniuje w mitochondriach, chloroplastach i bakteriach i jest powiązany z transportem elektronów.
Kompleksy I i IV transportu elektronów działają jak pompy protonowe. Te ulegają zmianom konformacyjnym, które pozwalają im pompować protony do przestrzeni międzybłonowej. W kompleksie IV dla każdej pary elektronów dwa protony są wypompowywane z membrany, a dwa kolejne pozostają w matrycy, tworząc HdwaLUB.
Ubichinon w kompleksie III przyjmuje protony z kompleksów I i II i uwalnia je na zewnątrz błony. Każdy kompleks I i III umożliwia przejście czterech protonów na każdą parę transportowanych elektronów.
Macierz mitochondrialna ma niskie stężenie protonów i ujemny potencjał elektryczny, natomiast w przestrzeni międzybłonowej występują warunki odwrotne. Przepływ protonów przez tę membranę reprezentuje gradient elektrochemiczny, który magazynuje energię niezbędną (± 5 kcal / mol na proton) do syntezy ATP..
Enzym syntetaza ATP jest piątym kompleksem biorącym udział w fosforylacji oksydacyjnej. Odpowiada za wykorzystanie energii gradientu elektrochemicznego do wytworzenia ATP.
To białko transbłonowe składa się z dwóch składników: F.0 i F.1. Składnik F.0 umożliwia powrót protonów do macierzy mitochondrialnej funkcjonującej jako kanał i F1 katalizuje syntezę ATP przez ADP i P.ja, wykorzystując energię wspomnianego powrotu.
Proces syntezy ATP wymaga zmiany strukturalnej w F.1 i montaż elementów F.0 i F.1. Translokacja protonu przez F0 powoduje zmiany konformacyjne w trzech podjednostkach F1, pozwalając mu działać jako silnik rotacyjny, kierując powstawaniem ATP.
Podjednostka odpowiedzialna za wiązanie ADP z Pja przechodzi ze stanu słabego (L) do stanu aktywnego (T). Kiedy tworzy się ATP, druga podjednostka przechodzi w stan otwarty (O), który umożliwia uwolnienie tej cząsteczki. Po uwolnieniu ATP ta podjednostka przechodzi ze stanu otwartego do stanu nieaktywnego (L).
Cząsteczki ADP i P.ja wiąże się z podjednostką, która przeszła ze stanu O do stanu L..
Łańcuch transportu elektronów i fosforylacja wytwarzają cząsteczki ATP. Utlenianie NADH daje około 52,12 kcal / mol (218 kJ / mol) darmowej energii.
Ogólna reakcja utleniania NADH jest następująca:
NADH + 1⁄2 Odwa +H.+ ↔ H.dwaO + NAD+
Transfer elektronów z NADH i FADHdwa zachodzi poprzez różne kompleksy, pozwalając na podzielenie swobodnej zmiany energii ΔG ° na mniejsze „pakiety” energii, które są sprzężone z syntezą ATP.
Utlenianie jednej cząsteczki NADH powoduje syntezę trzech cząsteczek ATP. Podczas utleniania cząsteczki FADHdwa par do syntezy dwóch ATP.
Te koenzymy pochodzą z procesów glikolizy i kwasu cytrynowego. Każda zdegradowana cząsteczka glukozy ostatecznie wytwarza 36 lub 38 cząsteczek ATP, w zależności od lokalizacji komórek. 36 ATP jest wytwarzane w mózgu i mięśniach szkieletowych, podczas gdy 38 ATP jest wytwarzane w tkance mięśniowej..
Wszystkie organizmy, jednokomórkowe i wielokomórkowe, potrzebują minimalnej ilości energii w swoich komórkach do przeprowadzania zachodzących w nich procesów, a tym samym do utrzymania funkcji życiowych w całym organizmie..
Procesy metaboliczne wymagają energii. Większość energii użytkowej uzyskuje się z rozkładu węglowodanów i tłuszczów. Energia ta pochodzi z procesu fosforylacji oksydacyjnej.
Stopień wykorzystania ATP w komórkach kontroluje jego syntezę, az kolei, ze względu na sprzężenie fosforylacji oksydacyjnej z łańcuchem transportu elektronów, na ogół reguluje również szybkość transportu elektronów..
Fosforylacja oksydacyjna podlega ścisłej kontroli, która zapewnia, że ATP nie jest wytwarzane szybciej niż jest zużywane. Istnieją pewne etapy procesu transportu elektronów i sprzężonej fosforylacji, które regulują tempo produkcji energii..
Główne szlaki produkcji energii (komórkowe ATP) to glikoliza, cykl kwasu cytrynowego i fosforylacja oksydacyjna. Skoordynowana kontrola tych trzech procesów reguluje syntezę ATP.
Kontrola fosforylacji przez stosunek masowego działania ATP zależy od dokładnego udziału elektronów w łańcuchu transportowym. To z kolei zależy od [NADH] / [NAD+], który jest podwyższony przez działanie glikolizy i cykl kwasu cytrynowego.
Ta skoordynowana kontrola jest przeprowadzana poprzez regulację punktów kontrolnych glikolizy (PFK hamowany przez cytrynian) i cyklu kwasu cytrynowego (dehydrogenaza pirogronianowa, taśma cytrynianowa, dehydrogenaza izocytrynianowa i dehydrogenaza α-ketoglutaranu)..
Kompleks IV (oksydaza cytochromu c) jest enzymem regulowanym przez jeden z jego substratów, czyli jego aktywność jest kontrolowana przez zredukowany cytochrom c (cdwa+), co z kolei jest w równowadze ze stosunkiem stężeń między [NADH] / [NAD+] i stosunek masy działania [ATP] / [ADP] + [Pja].
Im wyższy stosunek [NADH] / [NAD+] i obniż [ATP] / [ADP] + [Pja], tym wyższe stężenie cytochromu [cdwa+] i im większa aktywność kompleksu IV. Jest to interpretowane na przykład, jeśli porównamy organizmy o różnych aktywnościach odpoczynku i wysokiej aktywności.
U osoby o dużej aktywności fizycznej zużycie ATP, a tym samym jego hydroliza do ADP + Pja będzie bardzo duża, generując różnicę w stosunku masowego działania powodującą wzrost [cdwa+], a zatem wzrost syntezy ATP. U osoby w stanie spoczynku występuje sytuacja odwrotna.
Ostatecznie tempo fosforylacji oksydacyjnej rośnie wraz ze stężeniem ADP w mitochondriach. To stężenie zależy od translokatorów ADP-ATP odpowiedzialnych za transport nukleotydów adeninowych i P.ja od cytozolu do macierzy mitochondrialnej.
Na fosforylację oksydacyjną wpływają pewne czynniki chemiczne, które pozwalają na kontynuację transportu elektronów bez występowania fosforylacji ADP, oddzielając produkcję i konserwację energii.
Środki te stymulują tempo zużycia tlenu przez mitochondria pod nieobecność ADP, powodując również wzrost hydrolizy ATP. Działają poprzez wyeliminowanie pośrednika lub przerwanie stanu energetycznego łańcucha transportu elektronów.
2,4-dinitrofenol, słaby kwas, który przechodzi przez błony mitochondrialne, jest odpowiedzialny za rozpraszanie gradientu protonów, ponieważ wiążą się z nimi po stronie kwasowej i uwalniają je po stronie zasadowej.
Związek ten był stosowany jako „pigułka dietetyczna”, ponieważ stwierdzono, że powoduje przyspieszenie oddychania, a tym samym wzrost tempa metabolizmu i związaną z tym utratę wagi. Jednak wykazano, że jego negatywny wpływ może nawet spowodować śmierć.
Rozpraszanie gradientu protonów wytwarza ciepło. Komórki w brązowej tkance tłuszczowej wytwarzają ciepło za pomocą kontrolowanego hormonalnie rozprzęgania. Hibernujące ssaki i noworodki, którym brakuje włosów, składają się z tej tkanki, która służy jako rodzaj koca termicznego..
Zarówno związki, jak i środki hamujące zapobiegają zużyciu O.dwa (transport elektronów), takie jak powiązana fosforylacja oksydacyjna. Czynniki te zapobiegają tworzeniu się ATP poprzez wykorzystanie energii wytwarzanej w transporcie elektronicznym. Dlatego łańcuch transportowy zatrzymuje się, ponieważ wspomniane zużycie energii nie jest dostępne..
Antybiotyk oligomycyna działa jako inhibitor fosforylacji u wielu bakterii, zapobiegając stymulacji syntezy ADP do ATP.
Istnieją również środki jonoforowe, które tworzą rozpuszczalne w tłuszczach kompleksy z kationami, takimi jak K+ i Na+, i przechodzą przez błonę mitochondrialną z tymi kationami. Mitochondria wykorzystują następnie energię wytwarzaną w transporcie elektronów do pompowania kationów zamiast syntetyzowania ATP..


Jeszcze bez komentarzy