
Plik dihybrydyzm, W genetyce definiuje równoczesne badanie dwóch różnych cech dziedzicznych, a co za tym idzie, tych, których manifestacja zależy od dwóch różnych genów, nawet jeśli jest to ta sama postać.
Siedem cech, które analizował Mendel, było dla niego użytecznych w przedstawianiu teorii dziedziczenia postaci, ponieważ, między innymi, geny odpowiedzialne za ich manifestację miały kontrastujące allele, których fenotyp był łatwy do analizy, a każdy z nich określał wyrażenie jako pojedynczy postać.
Oznacza to, że były to cechy monogeniczne, których stan hybrydowy (monohybrydy) pozwolił określić relacje dominacji / recesywności między allelami tego pojedynczego genu..
Kiedy Mendel przeanalizował wspólne dziedziczenie dwóch różnych postaci, postąpił tak samo, jak z pojedynczymi postaciami. Otrzymał podwójne hybrydy (dihybrydy), które pozwoliły mu sprawdzić:
Teraz wiemy, że dziedziczenie znaków jest nieco bardziej złożone niż to, co zaobserwował Mendel, ale także, że w swoich podstawach Mendel był całkowicie poprawny..
Późniejszy rozwój genetyki pozwolił wykazać, że krzyżówki dwuhybrydowe i ich analiza (dihybrydyzm), jak początkowo był w stanie wykazać Bateson, mogą być niewyczerpanym źródłem odkryć w tej potężnej i rodzącej się nauce XX wieku..
Dzięki ich sprytnemu wykorzystaniu byli w stanie dać genetykowi nieco jaśniejszy obraz zachowania i natury genów..
Indeks artykułów
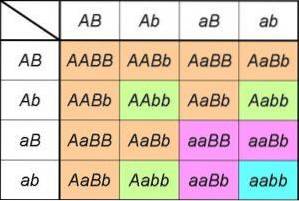
Jeśli przeanalizujemy produkty krzyża monohybrydowego Aa X Aa, możemy zauważyć, że jest to równoznaczne z opracowaniem niezwykłego produktu (DO+do)dwa= AA + 2Aa + aa.
Wyrażenie po lewej stronie obejmuje dwa typy gamet, które może wytworzyć jedno z rodziców heterozygotycznych pod względem genu DO/do; poprzez podniesienie do kwadratu wskazujemy, że oboje rodzice mają identyczną budowę dla badanego genu [to znaczy jest to krzyżówka monohybrydowa (DO+do) X (DO+do)].
Wyrażenie po prawej stronie podaje nam genotypy (a zatem fenotypy są wywnioskowane) i oczekiwane proporcje wynikające ze skrzyżowania.
Stąd bezpośrednio obserwujemy proporcje genotypowe wyprowadzone z pierwszego prawa (1: 2: 1), a także wyjaśniane przez nie proporcje fenotypowe (1AA+dwaAa= 3DO_ za każde 1aa, lub stosunek fenotypowy 3: 1).
Jeśli teraz rozważymy krzyż, aby przeanalizować dziedziczenie genu b, wyrażenia i proporcje będą takie same; w rzeczywistości tak będzie w przypadku każdego genu. Zatem w krzyżówce dwuhybrydowej faktycznie mamy rozwój produktów (DO+do)dwa X (b+b)dwa.
Albo co jest tym samym, jeśli krzyżówka dwuhybrydowa obejmuje dwa geny, które uczestniczą w dziedziczeniu dwóch niepowiązanych ze sobą postaci, proporcje fenotypowe będą takie, jak przewiduje drugie prawo: (3DO_: 1aa) X (3b_: 1kochanie) = 9DO_b_: 3DO_kochanie: 3aaB_: 1aabb).
Te oczywiście wywodzą się z uporządkowanych proporcji genotypu 4: 2: 2: 2: 2: 1: 1: 1: 1 wynikających z iloczynu (DO+do)dwa X (b+b)dwa= (AA + 2Aa + aa) X (nocleg ze śniadaniem + dwaNocleg ze śniadaniem + kochanie).
Zachęcamy do samodzielnego przekonania się i przeanalizowania teraz, co się dzieje, gdy proporcje fenotypowe 9: 3: 3: 1 krzyżówki dwuhybrydowej „odbiegają” od tych jasnych i przewidywalnych zależności matematycznych, które wyjaśniają niezależne dziedziczenie dwóch zakodowanych znaków. By różne geny.
Istnieją dwa główne sposoby, w jakie krzyżówki dwuhybrydowe odbiegają od tego, co jest „oczekiwane”. Pierwsza to ta, w której analizujemy wspólne dziedziczenie dwóch różnych postaci, ale proporcje fenotypowe obserwowane u potomstwa dają wyraźną przewagę przejawianiu się fenotypów rodzicielskich..
Najprawdopodobniej jest to przypadek połączonych genów. Oznacza to, że dwa analizowane geny, chociaż znajdują się w różnych loci, są tak blisko siebie fizycznie, że mają tendencję do dziedziczenia razem i, oczywiście, nie są dystrybuowane niezależnie..
Druga okoliczność, również dość powszechna, wynika z faktu, że niewielka mniejszość cech dziedzicznych jest monogeniczna..
Wręcz przeciwnie, w przejawianiu większości cech dziedzicznych uczestniczą więcej niż dwa geny..
Z tego powodu zawsze jest możliwe, że interakcje genetyczne, które są ustalane między genami uczestniczącymi w przejawianiu się jednego charakteru, są złożone i wykraczają poza prosty związek dominacji lub recesywności, jak obserwuje się w związkach allelicznych typowych dla cech monogenicznych.
Na przykład, w przejawianiu się cechy, około czterech enzymów może uczestniczyć w określonej kolejności, dając początek końcowemu produktowi odpowiedzialnemu za fenotypową manifestację dzikiego fenotypu..
Analiza, która pozwala zidentyfikować liczbę genów z różnych loci, które uczestniczą w manifestacji cechy genetycznej, a także kolejność, w jakiej działają, nazywa się analizą epistazy i jest prawdopodobnie tą, która najczęściej definiuje to, co my nazywamy analizą genetyczną w jej najbardziej klasycznym sensie.
Na końcu tego posta przedstawiono proporcje fenotypowe obserwowane w najczęstszych przypadkach epistazy - i to tylko z uwzględnieniem krzyżówek dwuhybrydowych..
Zwiększając liczbę genów, które uczestniczą w przejawianiu się tej samej cechy, oczywiście zwiększa się złożoność interakcji genów i ich interpretacji..
Ponadto, co z kolei można przyjąć jako złotą zasadę prawidłowej diagnozy oddziaływań epistatycznych, można zweryfikować pojawienie się nowych fenotypów nieobecnych w pokoleniu rodzicielskim..
Wreszcie, oprócz umożliwienia analizy pojawiania się nowych fenotypów i ich proporcji, analiza epistazy pozwala również określić hierarchiczny porządek, w jakim różne geny i ich produkty muszą przejawiać się na danej ścieżce, aby uwzględnić związany z nimi fenotyp..
Najbardziej podstawowy lub wczesny gen manifestacyjny jest epistatyczny nad wszystkimi innymi, ponieważ bez jego produktu lub działania, na przykład, osoby poniżej niego nie będą w stanie wyrazić siebie, co w związku z tym będzie dla niego hipostatyczne..
Gen / produkt na trzecim miejscu w hierarchii będzie hipostatyczny w stosunku do pierwszych dwóch i epistatyczny w stosunku do wszystkich pozostałych pozostających na tym szlaku ekspresji genów..



Jeszcze bez komentarzy