Plik epistaza, w genetyce jest to badanie interakcji między różnymi genami, które kodują tę samą postać. Oznacza to, że jest to przejaw cechy, która powstaje w wyniku interakcji między allelami genów w różnych loci.
Kiedy mówimy o związkach, które ustanawiają allele tego samego genu, mamy na myśli związki alleliczne. To znaczy do alleli tego samego locus lub allelomorficznych alleli. Są to dobrze znane interakcje całkowitej dominacji, niepełnej dominacji, kodominacji i śmiertelności między allelami tego samego genu..
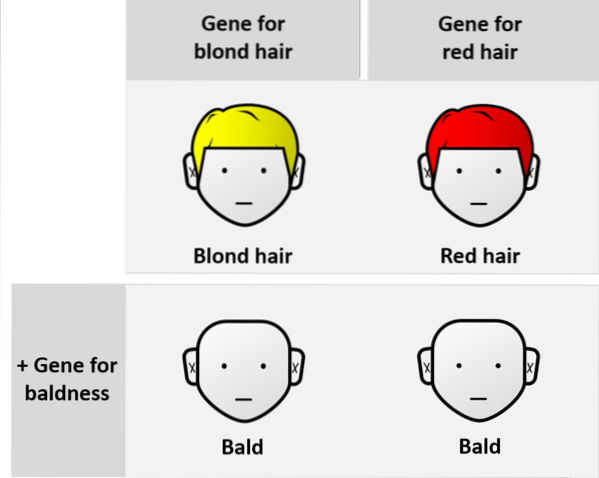
Przeciwnie, w relacjach między allelami różnych loci mówimy o allelach nie-allelomorficznych. Są to tak zwane interakcje genów, które w jakiś sposób są epistatyczne.
Epistasis umożliwia analizę, czy ekspresja jednego genu determinuje ekspresję innego. W takim przypadku taki gen byłby epistatyczny na drugim; drugi byłby hipostatyczny na pierwszym. Analiza epistazy pozwala również określić kolejność, w jakiej działają geny definiujące ten sam fenotyp.
Najprostsza epistaza analizuje, w jaki sposób dwa różne geny oddziałują na siebie, aby nadać ten sam fenotyp. Ale oczywiście może to być znacznie więcej genów.
W celu analizy prostej epistazy opieramy się na wariacjach proporcji klasycznych krzyżówek dwuhybrydowych. To znaczy do modyfikacji stosunku 9: 3: 3: 1 i do siebie.
Indeks artykułów
Proporcja ta wynika z połączenia analizy dziedziczenia dwóch różnych postaci. Oznacza to, że jest to produkt połączenia dwóch niezależnych segregacji fenotypowych (3: 1) X (3: 1).
Kiedy Mendel analizował, na przykład, wzrost rośliny lub kolor nasion, każdy znak segregował od 3 do 1. Kiedy analizował je razem, nawet jeśli były to dwie różne postacie, każdy z nich segregował od 3 do 1. Oznacza to, że były one rozdzielane niezależnie.
Jednak kiedy Mendel analizował postacie w parach, zaowocowały dobrze znanymi klasami fenotypowymi 9, 3, 3 i 1. Ale te klasy były sumami dwóch znaków. różne. I nigdy, żadna z postaci nie wpływała na to, jak manifestowała się druga.
Poprzednia była wyjaśnieniem klasycznej proporcji mendlowskiej. Dlatego nie jest to przypadek epistazy. Epistasis bada przypadki dziedziczenia o tym samym charakterze, zdeterminowane przez kilka genów.
Poprzedni przypadek, czyli drugie prawo Mendla, dotyczyło dziedziczenia dwóch różnych postaci. Te wyjaśnione później są prawdziwymi epistatycznymi proporcjami i dotyczą tylko allelomorficznych alleli..
Ten przypadek ma miejsce, gdy ta sama postać ma cztery różne objawy fenotypowe w stosunku 9: 3: 3: 1. Dlatego nie może to być interakcja alleliczna (monogeniczna), taka jak ta, która prowadzi do pojawienia się czterech różnych grup krwi w układzie ABO..
Weźmy jako przykład skrzyżowanie osoby heterozygotycznej z krwią typu A i osoby heterozygotycznej z krwią typu B. jaDOja X jabja. To dałoby nam stosunek osobników 1: 1: 1: 1 jaDOja (Typ A), jaDOjab (Typ AB), jabja (Typ B) e ii (Typ O).
Wręcz przeciwnie, prawdziwie dominujący podwójny związek epistatyczny (9: 3: 3: 1) jest obserwowany w postaci grzebieni koguta. Istnieją cztery klasy fenotypowe, ale w stosunku 9: 3: 3: 1.
W jego determinacji i przejawianiu uczestniczą dwa geny, nazwijmy je R Y P.. Niezależnie od alleli R Y P. wykazują całkowitą dominację nad allelami r Y p, odpowiednio.
Przejścia RrPp X RrPp możemy otrzymać klasy fenotypowe 9 R_P_, 3 R_pp, 3 rrP_ i 1 rrpp. Symbol „_” oznacza, że allel może być dominujący lub recesywny. Powiązany fenotyp pozostaje ten sam.
Klasa 9 R_P_ jest reprezentowana przez koguty z herbem orzecha włoskiego, 3 R_pp przez koguty z herbem róży. Koguty czubate będą należały do klasy 3 rrP_; te z klasy rrpp mają prosty grzebień.

W epistazie z podwójną dominacją każda klasa 3 wynika z efektu dominacji genu R lub P. Klasa 9 jest reprezentowana przez ten, w którym manifestują się allele dominujące zarówno R, jak i P. Wreszcie, w klasie 1 rrpp allele są nieobecne. obu genów.
W tej epistatycznej interakcji jeden gen nie tłumi manifestacji innego. Wręcz przeciwnie, oba geny kodują manifestację tej samej cechy, ale bez efektu addytywnego..
Dlatego obecność co najmniej jednego dominującego allelu któregokolwiek z dwóch genów z różnych loci pozwala na manifestację cechy w klasie 15. Brak dominujących alleli (klasa podwójnie recesywna) determinuje fenotyp klasy 1..
Produkty genów uczestniczą w przejawianiu koloru ziarna pszenicy DO mnie b. Oznacza to, że każdy z tych produktów (lub oba) może prowadzić do reakcji biochemicznej, która przekształca prekursor w pigment..
Jedyną klasą, która nie produkuje żadnego z nich, jest klasa 1 aabb. Dlatego klasy 9 A_B_, 3 A_bb i 3 aaB_ będą wytwarzać ziarna pigmentowane, a pozostała mniejszość nie będzie.
Tutaj znajdujemy przypadek dominującej delecji genu (hipostatycznej) z powodu obecności co najmniej jednego dominującego allelu drugiego (epistatycznego). Oznacza to, że formalnie rzecz biorąc, jeden gen tłumi działanie drugiego..
Jeśli jest to dominujące tłumienie D nad K, mielibyśmy ten sam fenotyp związany z klasami 9 D_K_, 3 D_kk i 1 ddkk. Klasa 3 ddK_ byłaby jedyną, która wykazywałaby nie tłumioną cechę.
Klasa podwójnie recesywna jest dodawana do klas 9 D_K_ i 3 D_kk, ponieważ nie wytwarza tego, co koduje hipostatyczny gen K. Nie dlatego, że jest stłumiony przez D, którego w każdym razie też nie ma, ale ponieważ nie wytwarza K.
Ten stosunek jest czasami nazywany epistazą dominującą i recesywną. Dominuje K. na D / d. Recesywna epistaza byłaby epistazą dd na K / k.
Na przykład kwiaty wiesiołka zawdzięczają swój kolor manifestacji dwóch genów. Gen K. który koduje produkcję pigmentu malwidyna i gen re które kody do usunięcia malvidin.
Tylko rośliny ddKK lub ddKk (tj. klasa 3 ddK_) będzie produkować malwidynę i będzie koloru niebieskiego. Każdy inny genotyp da początek roślinom o turkusowych kwiatach.
W tym przypadku obecność przynajmniej jednego dominującego allelu każdego genu w parze jest wymagana, aby postać się zamanifestowała. Powiedzmy, że to geny do Y P.. Oznacza to, że homozygotyczny recesywny stan jednego z genów pary (DC lub s) uniemożliwia manifestację postaci.
Innymi słowy, tylko klasa 9 C_P_ ma co najmniej jeden dominujący allel do i dominujący allel P.. Aby cecha się zamanifestowała, muszą być obecne funkcjonalne produkty obu genów.
Ta interakcja jest epistatyczna, ponieważ brak ekspresji jednego genu uniemożliwia manifestację drugiego genu. Jest podwójna, ponieważ prawdą jest również odwrotność.
Klasycznym przykładem ilustrującym ten przypadek są kwiaty grochu. Rośliny CCpp i rośliny ccPP mają białe kwiaty. Hybrydy CcPp krzyżówek między nimi przedstawiają fioletowe kwiaty.
Jeśli skrzyżowane zostaną dwie z tych roślin dwuhybrydowych, otrzymamy klasę 9 C-P_, która będzie miała fioletowe kwiaty. Klasy 3 C_pp, 3 ccP_ i ccpp będą białymi kwiatami.

Z proporcji zaproponowanych w drugim prawie Mendla mamy inne dodatkowe przypadki, które zasługują na wzmiankę..
Nie bez powodu nazywamy zmodyfikowaną epistazą recesywną o zmodyfikowanym stosunku 9: 4: 3. Kiedy gen jest homozygotyczny dla genu recesywnego, unika ekspresji drugiego genu - nawet jeśli jest dominujący.
Weźmy na przykład recesywną epistazę genotypu aa o tych z genu b. Klasa 9 to już rozpoznane 9 A_B_. Dla klasy 4, do klasy 1 aabb, należy dodać te z klasy 3 aaB_, o tym samym fenotypie. Klasa 3 byłaby klasą 3 A_bb.
W epistatycznej interakcji zduplikowanych genów obserwowany stosunek fenotypowy wynosi 9: 6: 1. Wszystkie osobniki klasy 9 A_B_ mają co najmniej jeden allel każdego genu DO lub b. Wszystkie mają ten sam fenotyp.
Wręcz przeciwnie, w klasach 3 A_bb i 3 aaBb występują tylko dominujące allele A lub B. W tym przypadku istnieje również jeden i ten sam fenotyp - ale różni się od pozostałych. Wreszcie, w klasie 1 aabb nie występuje żaden dominujący allel żadnego z genów i reprezentuje inny fenotyp.
Być może najbardziej zagmatwaną klasą jest dominująca epistaza, która wykazuje stosunek fenotypowy 12: 3: 1. Tutaj dominacja A (epistatyczna) nad B (hipostatyczna) powoduje, że klasa 9 A-B_ dołącza do klasy 3 A_bb.
Fenotyp B przejawia się tylko wtedy, gdy A nie występuje w klasie 3 aaB_. Podwójnie recesywny aabb klasy 1 nie przejawia się ani nie przejawia fenotypu związanego z genem A / a ani z genem B / b.
Inne epistatyczne współczynniki fenotypowe, które nie mają określonej nazwy, to 7: 6: 3, 3: 6: 3: 4 i 11: 5.


Jeszcze bez komentarzy