Plik ekologiczne prawo dziesięciny, prawo ekologiczne lub 10% podnosi sposób, w jaki energia przemieszcza się w jej pochodzeniu przez różne poziomy troficzne. Często twierdzi się również, że ta zasada jest po prostu bezpośrednią konsekwencją drugiej zasady termodynamiki.
Energia ekologiczna jest częścią ekologii, która dotyczy ilościowego określenia zależności, które nakreśliliśmy powyżej. Uważa się, że Raymond Lindemann (szczególnie w swojej przełomowej pracy z 1942 r.) Był tym, który stworzył podstawy tego kierunku studiów.
Jego praca koncentrowała się na pojęciach łańcucha pokarmowego i sieci oraz na kwantyfikacji wydajności transferu energii między różnymi poziomami troficznymi..
Lindemann rozpoczyna od padającego promieniowania słonecznego lub energii, którą otrzymuje społeczność, poprzez wychwytywanie dokonywane przez rośliny w procesie fotosyntezy i kontynuuje monitorowanie tego wychwytywania i jego późniejszego wykorzystania przez roślinożerców (konsumentów pierwotnych), następnie przez mięsożerców (konsumentów wtórnych), a na końcu przez rozkładających się.
Indeks artykułów
Po pionierskiej pracy Lindemanna przyjęto, że efektywność transferu troficznego wynosi około 10%; w rzeczywistości niektórzy ekologowie odnosili się do 10% prawa. Jednak od tego czasu pojawiło się wiele nieporozumień dotyczących tej kwestii..
Z pewnością nie ma prawa natury, które powoduje, że dokładnie jedna dziesiąta energii wchodzącej na jeden poziom troficzny jest przenoszona na następny..
Na przykład zestawienie badań troficznych (w środowiskach morskich i słodkowodnych) ujawniło, że skuteczność przenoszenia według poziomu troficznego wahała się między około 2 a 24%, chociaż średnia wynosiła 10,13%..
Zasadniczo, mającą zastosowanie zarówno do systemów wodnych, jak i lądowych, można powiedzieć, że drugorzędna produktywność zwierząt roślinożernych znajduje się zwykle w przybliżeniu, o rząd wielkości poniżej pierwotnej produktywności, na której jest oparta..
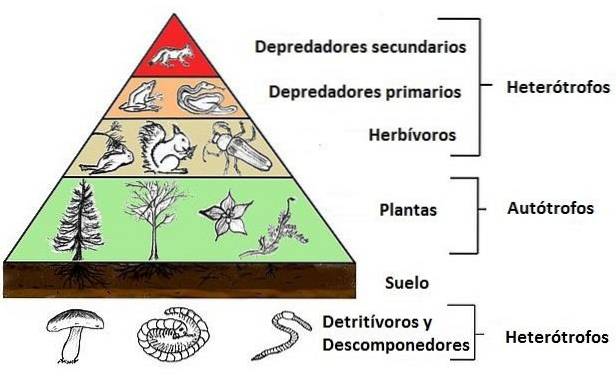
Często jest to spójna relacja, która jest utrzymywana we wszystkich systemach żerowania i która ma tendencję do przekształcania się w struktury typu piramidalnego, w których podstawę stanowią rośliny i na tej podstawie opiera się mniejsza, pierwotnych konsumentów, na których inny (jeszcze mniejsi) wtórni konsumenci siedzą.
Wszystkie żywe istoty wymagają materii i energii; materia dla budowy ich ciał i energii potrzebnej do pełnienia funkcji życiowych. Wymóg ten nie ogranicza się do pojedynczego organizmu, ale rozciąga się na wyższe poziomy organizacji biologicznej, które te osoby mogą spełnić..
Te poziomy organizacji to:
W ekosystemie społeczność i środowisko ustanawiają przepływy energii i materii.
Organizmy w ekosystemie są pogrupowane według „roli” lub „funkcji”, jaką pełnią w łańcuchu pokarmowym lub troficznym; tak mówimy o poziomach troficznych producentów, konsumentów i rozkładających się.
Z kolei każdy z tych poziomów troficznych oddziałuje ze środowiskiem fizykochemicznym, które zapewnia warunki do życia, a jednocześnie działa jako źródło i zlew energii i materii..
Najpierw musimy zdefiniować produktywność pierwotną, czyli tempo, w jakim biomasa jest produkowana na jednostkę powierzchni..
Zwykle jest wyrażana w jednostkach energii (dżule na metr kwadratowy dziennie) lub w jednostkach suchej masy organicznej (kilogramy na hektar na rok) lub jako węgiel (masa węgla w kg na metr kwadratowy na rok).
Ogólnie mówiąc, kiedy odnosimy się do całej energii związanej z fotosyntezą, zwykle nazywamy ją pierwotną produktywnością brutto (PPG).
Część z tego jest zużywana na oddychanie tych samych autotrofów (RA) i jest tracona w postaci ciepła. Pierwotną produkcję netto (PPN) uzyskuje się odejmując tę ilość od PPG (PPN = PPG-RA).
Ta pierwotna produkcja netto (PPN) jest ostatecznie dostępna do spożycia przez heterotrofy (są to bakterie, grzyby i pozostałe znane nam zwierzęta).
Produktywność wtórna (PS) jest definiowana jako tempo produkcji nowej biomasy przez organizmy heterotroficzne. W przeciwieństwie do roślin, bakterii heterotroficznych, grzybów i zwierząt nie mogą wytwarzać złożonych i bogatych w energię związków z prostych cząsteczek..
Swoją materię i energię czerpią zawsze z roślin, co mogą zrobić bezpośrednio, konsumując materiał roślinny lub pośrednio, żerując na innych heterotrofach..
W ten sposób rośliny lub organizmy fotosyntetyzujące w ogóle (zwane także producentami) stanowią pierwszy poziom troficzny w zbiorowisku; konsumenci pierwotni (ci, którzy żywią się producentami) stanowią drugi poziom troficzny, a konsumenci wtórni (zwani również mięsożernymi) stanowią trzeci poziom.
Proporcje pierwotnej produkcji netto, które przepływają wzdłuż każdej z możliwych ścieżek energii, ostatecznie zależą od wydajności transferu, to znaczy od sposobu, w jaki energia jest wykorzystywana i przekazywana z jednego poziomu na drugi..
Istnieją trzy kategorie efektywności transferu energii i dzięki ich dobrze zdefiniowanym możemy przewidzieć schemat przepływu energii na poziomach troficznych. Te kategorie to: efektywność zużycia (EC), efektywność asymilacji (EA) i wydajność produkcji (EP).
Zdefiniujmy teraz te trzy wspomniane kategorie.
Matematycznie możemy zdefiniować efektywność zużycia (EC) w następujący sposób:
EC =jan/P.n-1 × 100
Gdzie widzimy, że CE jest procentem całkowitej dostępnej produktywności (P.n-1), który jest skutecznie wchłaniany przez górny, ciągły przedział troficzny (jan).
Na przykład w przypadku konsumentów pierwotnych w systemie wypasu EC jest procentem (wyrażonym w jednostkach energii i na jednostkę czasu) spożycia PPN przez zwierzęta roślinożerne.
Gdybyśmy odnosili się do konsumentów wtórnych, byłoby to równoważne procentowi produktywności roślinożerców konsumowanej przez drapieżniki. Reszta ginie niezjedzona i wchodzi w łańcuch rozkładu.
Z drugiej strony efektywność asymilacji wyraża się następująco:
EA =DOn/jan × 100
Ponownie odnosimy się do procentu, ale tym razem do części energii pochodzącej z pożywienia i wchłoniętej przez konsumenta w komorze troficznej (jan) i jest przyswajane przez układ pokarmowy (DOn).
Ta energia będzie dostępna dla wzrostu i wykonania pracy. Pozostała część (część nie zasymilowana) jest tracona z kałem, a następnie wchodzi na poziom troficzny rozkładających się.
Wreszcie wydajność produkcji (EP) jest wyrażona jako:
EP = Pn/DOn × 100
co też jest procentem, ale w tym przypadku odnosimy się do energii asymilowanej (DOn), która kończy się włączeniem do nowej biomasy (P.n). Cała niezasymilowana pozostałość energii jest tracona w postaci ciepła podczas oddychania.
Produkty takie jak wydzieliny i / lub wydaliny (bogate w energię), które uczestniczyły w procesach metabolicznych, można uznać za produkcję, P.n, i są dostępne, jak zwłoki, dla rozkładających się.
Po zdefiniowaniu tych trzech ważnych kategorii, możemy teraz zadać sobie pytanie o „globalną efektywność transferu” z jednego poziomu troficznego na następny, którą po prostu daje iloczyn wyżej wspomnianych sprawności (EC x EA x EP).
Wyrażając kolokwialnie, możemy powiedzieć, że efektywność poziomu jest podana przez to, co można skutecznie połknąć, co jest następnie przyswajane i ostatecznie włączane do nowej biomasy.
Produktywność roślinożerców jest zawsze niższa niż roślin, którymi się żywią. Moglibyśmy wtedy zadać sobie pytanie: Gdzie idzie marnowana energia?
Aby odpowiedzieć na to pytanie, musimy zwrócić uwagę na następujące fakty:
Dzieje się tak z dwóch podstawowych powodów: Po pierwsze, ponieważ nie ma procesu konwersji energii, który byłby w 100% wydajny. Oznacza to, że podczas konwersji zawsze występuje strata w postaci ciepła, co jest w pełni zgodne z Drugą zasadą termodynamiki..
Po drugie, ponieważ zwierzęta muszą wykonywać pracę, która wymaga wydatkowania energii, a to z kolei pociąga za sobą nowe straty w postaci ciepła..
Wzorce te występują na wszystkich poziomach troficznych i jak przewiduje Drugie Prawo Termodynamiki, część energii, którą próbujemy przenieść z jednego poziomu na drugi, jest zawsze rozpraszana w postaci bezużytecznego ciepła..


Jeszcze bez komentarzy