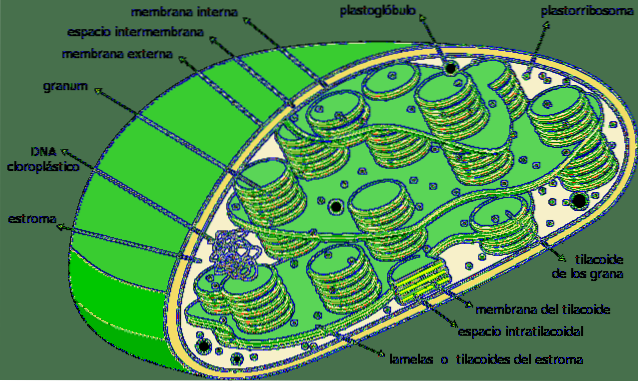
Plik tylakoidy to przedziały w postaci płaskich woreczków zlokalizowane w chloroplastach w komórkach roślinnych roślin, sinicach i algach. Zazwyczaj są zorganizowane w strukturę zwaną grana -plural granum- i wygląda jak stos monet.
Tylakoidy są uważane za trzeci system błonowy chloroplastów, poza wewnętrzną i zewnętrzną błoną wspomnianych organelli. Membrana tej struktury oddziela wnętrze tylakoidu od zrębu chloroplastu i zawiera szereg pigmentów i białek zaangażowanych w szlaki metaboliczne..
W tylakoidach zachodzą reakcje biochemiczne niezbędne do fotosyntezy, procesu, w którym rośliny pobierają światło słoneczne i przekształcają je w węglowodany. W szczególności mają one niezbędną maszynerię zakotwiczoną w membranie, aby przeprowadzić fazę zależną od słońca, w której światło jest uwięzione i przekształcone w energię (ATP) i NADPH..
Indeks artykułów
Tylakoidy są wewnętrznym trójwymiarowym błoniastym systemem chloroplastów. W pełni dojrzałe chloroplasty mają od 40 do 60 ułożonych w stos ziaren o średnicy od 0,3 do 0,6 µm..
Liczba tylakoidów, które tworzą kiełki, jest bardzo zróżnicowana: od mniej niż 10 woreczków u roślin wystawionych na dostateczne światło słoneczne do ponad 100 tylakoidów u roślin żyjących w skrajnie zacienionym środowisku..
Ułożone w stos tylakoidy są połączone ze sobą, tworząc ciągłą komorę wewnątrz chloroplastu. Wnętrze tylakoida to dość obszerny przedział o charakterze wodnistym..
Błona tylakoidów jest niezbędna do fotosyntezy, gdyż tam zachodzi pierwszy etap tego procesu.
Tylakoidy są dominującymi strukturami w pełni dojrzałego chloroplastu. Jeśli chloroplast jest wizualizowany w tradycyjnym mikroskopie świetlnym, można zaobserwować niektóre gatunki ziaren.
To są stosy tylakoidów; Z tego powodu pierwsi obserwatorzy tych struktur nazwali je „grana”.
Za pomocą mikroskopu elektronowego można było powiększyć obraz i wyciągnięto wniosek, że natura tych ziaren była w rzeczywistości ułożonymi tylakoidami.
Tworzenie i struktura błony tylakoidów zależy od tworzenia chloroplastu z jeszcze niezróżnicowanego plastydu, znanego jako protoplastyd. Obecność światła stymuluje konwersję do chloroplastów, a później tworzenie się spiętrzonych tylakoidów.
W chloroplastach i sinicach błona tylakoidów nie styka się z wewnętrzną częścią błony komórkowej. Jednak tworzenie błony tylakoidów rozpoczyna się od wpuszczenia błony wewnętrznej..
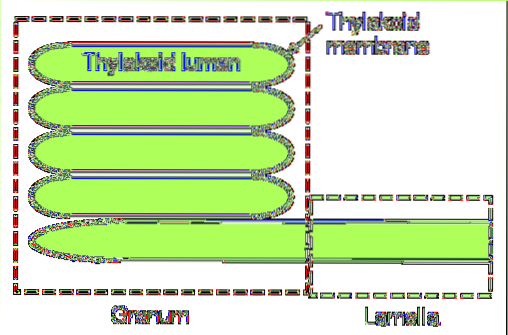
W sinicach i niektórych gatunkach glonów tylakoidy składają się z pojedynczej warstwy blaszek. Natomiast w dojrzałych chloroplastach występuje bardziej złożony system..
W tej ostatniej grupie można wyróżnić dwie zasadnicze części: grana i blaszkę zrębu. Pierwsza składa się z małych ułożonych w stos krążków, a druga jest odpowiedzialna za połączenie tych stosów ze sobą, tworząc ciągłą strukturę: światło tylakoidu..
Lipidy tworzące błonę są wysoce wyspecjalizowane i składają się w prawie 80% z galaktozylodiacyloglicerolu: monogalaktozylodiacyloglicerolu i digalaktozylodiacyloglicerolu. Te galaktolipidy mają wysoce nienasycone łańcuchy, typowe dla tylakoidów..
Podobnie błona tylakoidów zawiera mniej lipidów, takich jak fosfatydyloglicerol. Wspomniane lipidy nie są rozmieszczone równomiernie w obu warstwach membrany; istnieje pewien stopień asymetrii, który wydaje się przyczyniać się do funkcjonowania konstrukcji.
Fotoukłady I i II są dominującymi składnikami białek w tej błonie. Są one związane z kompleksem cytochromu b6Syntetaza F i ATP.
Stwierdzono, że większość elementów fotosystemu II znajduje się w ułożonych w stos błonach szkarłatnych, podczas gdy fotosystem I znajduje się głównie w nie ułożonych błonach tylakoidów. Oznacza to, że istnieje fizyczna separacja między dwoma fotosystemami..
Kompleksy te obejmują integralne białka błonowe, białka obwodowe, kofaktory i różne pigmenty..
Wnętrze tylakoidów składa się z gęstej, wodnistej substancji, której skład różni się od zrębu. Uczestniczy w fotofosforylacji, przechowując protony, które będą generować siłę napędową protonów do syntezy ATP. W tym procesie pH światła może osiągnąć 4.
W proteomie światła organizmu modelowego Arabidopsis thaliana zidentyfikowano ponad 80 białek, ale ich funkcje nie zostały w pełni wyjaśnione.
Białka światła są zaangażowane w regulację biogenezy tylakoidów oraz w aktywność i obrót białek tworzących kompleksy fotosyntetyczne, zwłaszcza fotosystemu II i dehydrogenazy NAD (P) H..
W tylakoidach rozpoczyna się niezbędny dla roślin proces fotosyntezy. Błona, która oddziela je od zrębu chloroplastów, ma całą maszynerię enzymatyczną niezbędną do zajścia reakcji fotosyntezy..
Fotosyntezę można podzielić na dwa główne etapy: reakcje na światło i reakcje w ciemności.
Jak sama nazwa wskazuje, reakcje należące do pierwszej grupy mogą zachodzić tylko w obecności światła, podczas gdy reakcje z drugiej grupy mogą zachodzić ze światłem lub bez niego. Zwróć uwagę, że otoczenie nie musi być „ciemne”, jest jedynie niezależne od światła.
Pierwsza grupa reakcji, „lekka”, zachodzi w tylakoidzie i można ją podsumować następująco: lekka + chlorofil + 12 HdwaO + 12 NADP+ + 18 ADP + 18 Pja à 6 Odwa + 12 NADPH + 18 ATP.
Druga grupa reakcji zachodzi w zrębie chloroplastowym i pobiera ATP i NADPH syntetyzowane w pierwszym etapie w celu redukcji węgla z dwutlenku węgla do glukozy (C6H.12LUB6). Drugi etap można podsumować jako: 12 NADPH + 18 ATP + 6 COdwa à C6H.12LUB6 + 12 NADP+ + 18 ADP + 18 Pja + 6 godzdwaLUB.
Reakcje świetlne obejmują szereg struktur znanych jako fotosystemy, które znajdują się w błonie tylakoidów i zawierają około 300 cząsteczek pigmentu, w tym chlorofilu..
Istnieją dwa typy fotosystemów: pierwszy ma maksymalną wartość szczytową pochłaniania światła 700 nanometrów i jest znany jako P.700, podczas gdy druga nazywa się P680. Obie są osadzone w błonie tylakoidów.
Proces rozpoczyna się, gdy jeden z pigmentów zaabsorbuje foton, który „odbija się” w kierunku innych pigmentów. Kiedy cząsteczka chlorofilu pochłania światło, jeden elektron wyskakuje, a inna cząsteczka go pochłania. Cząsteczka, która straciła elektron, jest teraz utleniona i ma ładunek ujemny.
P680 wychwytuje energię świetlną z chlorofilu a. W tym fotoukładzie elektron jest wrzucany do systemu o wyższej energii niż pierwotny akceptor elektronów..
Ten elektron wpada do fotosystemu I, przechodząc przez łańcuch transportu elektronów. Ten system reakcji utleniania i redukcji odpowiada za przenoszenie protonów i elektronów z jednej cząsteczki do drugiej..
Innymi słowy, następuje przepływ elektronów z wody do fotosystemu II, fotosystemu I i NADPH..
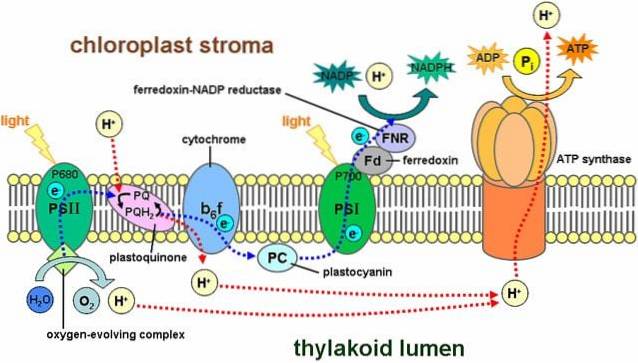
Część protonów generowanych przez ten układ reakcyjny znajduje się wewnątrz tylakoidów (zwanych także światłem tylakoidów), tworząc gradient chemiczny, który generuje siłę napędową protonów..
Protony przemieszczają się z przestrzeni tylakoidów do zrębu, zgodnie z gradientem elektrochemicznym; to znaczy wychodzą z tylakoidów.
Jednak protony nie przechodzą przez żadne miejsce w błonie, muszą to robić przez złożony układ enzymatyczny zwany syntetazą ATP.
Ten ruch protonów w kierunku zrębu powoduje powstawanie ATP wychodząc z ADP, procesu analogicznego do tego, który zachodzi w mitochondriach. Synteza ATP za pomocą światła nazywana jest fotofosforylacją.
Te wymienione etapy zachodzą jednocześnie: chlorofil z fotosystemu II traci elektron i musi go zastąpić elektronem z rozpadu cząsteczki wody; fotosystem I wyłapuje światło, utlenia się i uwalnia elektron uwięziony przez NADP+.
Utracony elektron z fotosystemu I zostaje zastąpiony elektronem wynikającym z fotosystemu II. Związki te zostaną wykorzystane w kolejnych reakcjach wiązania węgla w cyklu Calvina..
Ewolucja fotosyntezy jako procesu uwalniania tlenu umożliwiła życie, jakie znamy.
Sugeruje się, że fotosynteza rozwinęła się kilka miliardów lat temu u przodka, który dał początek obecnym cyjanobakteriom, z anoksycznego kompleksu fotosyntetycznego.
Proponuje się, że ewolucji fotosyntezy towarzyszyły dwa niezbędne wydarzenia: powstanie fotosystemu P.680 oraz geneza wewnętrznego systemu membranowego, bez połączenia z błoną komórkową.
Istnieje białko zwane Vipp1, niezbędne do tworzenia tylakoidów. Rzeczywiście, białko to występuje w roślinach, algach i sinicach, ale nie występuje w bakteriach, które przeprowadzają fotosyntezę beztlenową..
Uważa się, że gen ten mógł powstać w wyniku duplikacji genu u potencjalnego przodka cyjanobakterii. Jest tylko jeden przypadek cyjanobakterii, które są zdolne do fotosyntezy z tlenem i nie mają tylakoidów: gatunek Gloeobacter violaceus.


Jeszcze bez komentarzy