
Pojęcie podejście pozapiramidowe lub układ pozapiramidowy (SEP) powstał w wyniku badań anatomicznych i fizjologicznych, zmierzających do zrozumienia, w jaki sposób ośrodkowy układ nerwowy kontroluje aktywność mięśni szkieletowych, tak aby organizm przyjął odpowiednią postawę i wytworzył dobrowolną ruchy.
W tym procesie odkryto, że kontrola aktywności mięśni wymaga kontroli neuronów ruchowych przedniego rogu rdzenia kręgowego, jedynego połączenia między ośrodkowym układem nerwowym a włóknami mięśni szkieletowych, i że kontrolę tę sprawują projekcje nerwowe. z ośrodków mózgowych.
Wśród tych projekcji ważną ścieżkę tworzą niektóre aksony, które powstają w obszarach motorycznych kory mózgowej i schodzą bezpośrednio, to znaczy bez łusek, do rdzenia kręgowego, łącząc się, przechodząc przez rdzeń przedłużony, w niektórych wypukłościach które ze względu na swój kształt nazwano „piramidami”.
Ten przewód nazywano „piramidalnym” lub „korowo-rdzeniowym” i był zaangażowany w kontrolę drobnych i zręcznych ruchów wykonywanych przez dystalne części kończyn, podczas gdy istnienie struktur z funkcją motoryczną było rozpoznawane, ale nie uwzględnione. w ten sposób (extra).
Termin „pozapiramidowy układ ruchowy”, już przestarzały z fizjologicznego punktu widzenia, jest nadal używany w żargonie klinicznym w odniesieniu do tych struktur mózgu i pnia mózgu, które współpracują w zakresie kontroli motorycznej, ale nie są częścią układu piramidowego ani bezpośredniego połączenia korowo-rdzeniowego.
Indeks artykułów
Szlak pozapiramidowy można opisać jako zorganizowany w dwie grupy komponentów: jedna składałaby się z zestawu jąder pnia mózgu i jego projekcji w kierunku rdzenia kręgowego, a druga składałaby się z jąder podkorowych znanych jako jądra lub zwoje podstawy..
W pniu mózgu znajdują się grupy neuronów, których aksony wystają w kierunku istoty szarej rdzenia kręgowego i które zostały opisane jako zorganizowane w dwa układy: jeden przyśrodkowy i drugi boczny..
Układ przyśrodkowy tworzą drogi przedsionkowo-rdzeniowe, siateczkowo-rdzeniowe i tektospinalowe, które schodzą przez brzuszne sznury rdzenia i wywierają kontrolę nad mięśniami osiowymi lub tułowia, oprócz mięśni proksymalnych kończyn zaangażowanych w postawę ciała..
Najważniejszym elementem układu bocznego jest przewód rubrospinalowy, którego aksony wystają z czerwonego jądra śródmózgowia, schodzą przez boczny sznurek i wpływają na neurony ruchowe, które kontrolują dystalne mięśnie kończyn..
Z powyższego można wywnioskować, że układ przyśrodkowy współpracuje przy podstawowych korektach postawy, niezbędnych do dobrowolnej aktywności ruchowej, podczas gdy układ boczny zajmuje się, wraz z bezpośrednią drogą korowo-rdzeniową, ruchami kończyn, których celem jest np. sięganie i manipulowanie przedmiotami.
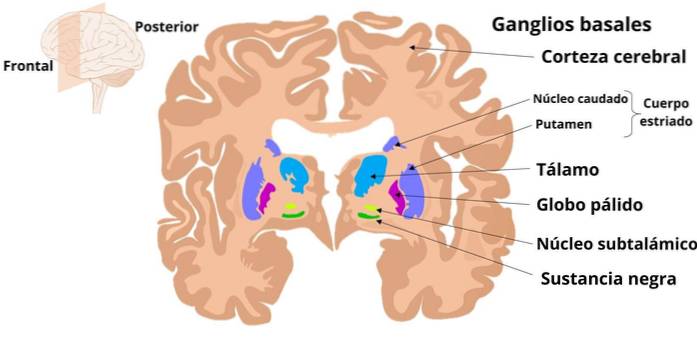
Zwoje podstawy to podkorowe struktury neuronalne, które biorą udział w przetwarzaniu informacji motorycznych, takich jak planowanie i programowanie złożonych, zręcznych ruchów, i których zmiany dają objawy kliniczne, które są pogrupowane w zespoły zwane „pozapiramidowymi”.
Ganglia obejmują prążkowie, które składa się ze skorupy i jądra ogoniastego; blada kula ziemska, która ma część zewnętrzną (GPe) i część wewnętrzną (GPi); istota czarna, zorganizowana w część zwartą (SNc) i część siatkową (SNr) oraz jądro podwzgórza lub Lewisa.
Struktury te działają poprzez otrzymywanie informacji głównie z różnych regionów kory mózgowej; informacja, która aktywuje obwody wewnętrzne, które wpływają na wyjściową aktywność neuronalną, która powraca, poprzez część motoryczną wzgórza, do kory mózgowej.
Informacje o zwojach przedostają się przez prążkowie (ogoniaste i skorupa). Stamtąd rozpoczynają się ścieżki, które łączą się z jądrami wyjściowymi, którymi są GPi i SNr, których aksony przechodzą do jąder przednio-wentylacyjnych i brzuszno-bocznych wzgórza, które z kolei wystają do kory.
Poszczególne etapy obwodu pokrywają neurony należące do określonego układu neurochemicznego, które mogą mieć działanie hamujące lub pobudzające. Połączenia korowo-prążkowane, włókna wzgórzowo-korowe i podwzgórze uwalniają glutaminian i działają pobudzająco.
Neurony, których aksony wychodzą z prążkowia, wykorzystują kwas gamma-aminomasłowy (GABA) jako główny neuroprzekaźnik i działają hamująco. Istnieją dwie subpopulacje: jedna syntetyzuje substancję P jako kotransmiter [GABA (+ Subst. P)], a druga enkefalina [GABA (+ Encef.)].
Neurony GABA (+ Sust. P) mają receptory dopaminy D1 i są pobudzane przez dopaminę (DA); ustanawiają również bezpośrednie połączenie hamujące z ujściami zwojów podstawy (GPi i SNr), które są również GABAergiczne, ale „+ dynorfinowe” i hamują glutaminergiczne komórki projekcji wzgórzowo-korowej.
GABA (+ Encef.) Neurony mają receptory dopaminy D2 i są hamowane przez dopaminę. Ustanawiają pośrednie połączenie pobudzające z wyjściami (GPi i SNr), ponieważ projektują do GPe hamując jego neurony GABAergiczne, które hamują neurony glutaminergiczne jądra podwzgórza, których funkcją jest aktywacja wyjść (GPi i SNr).
Zwarta część istoty czarnej (SNc) ma neurony dopaminergiczne (DA), które łączą się z prążkowiem tworząc połączenia, jak już wspomniano, pobudzające D1 na komórkach GABA (+ Sub. P) i hamujące D2 na komórkach GABA (+ Encef.).
Następnie, zgodnie z powyższym, aktywacja ścieżki bezpośredniej kończy się zahamowaniem wyjść zwojów podstawnych i uwolnieniem aktywności w połączeniach wzgórzowo-korowych, podczas gdy aktywacja ścieżki pośredniej aktywuje wyjścia i zmniejsza aktywność wzgórzową. -korowy.
Chociaż interakcje i dokładne wspólne funkcjonowanie właśnie rozważanych ścieżek bezpośrednich i pośrednich nie zostały wyjaśnione, opisana anatomiczna i neurochemiczna organizacja pomaga nam zrozumieć, przynajmniej częściowo, niektóre stany patologiczne wynikające z dysfunkcji zwojów podstawy..
Chociaż procesy patologiczne, które osiedlają się w zwojach podstawy mózgu, mają zróżnicowany charakter i wpływają nie tylko na określone funkcje motoryczne, ale także funkcje poznawcze, asocjacyjne i emocjonalne, na obrazach klinicznych zmiany motoryczne zajmują czołowe miejsce i większość badań skupiała się na nich..
Zaburzenia ruchowe typowe dla dysfunkcji zwojów podstawy można podzielić na jedną z trzech grup, a mianowicie:
- Hiperkinezje, takie jak choroba Huntingtona lub pląsawica i hemibalizm.
- Hipokinezie, takie jak choroba Parkinsona.
- Dystonie, takie jak atetoza.
Ogólnie można powiedzieć, że zaburzenia hiperkinetyczne, charakteryzujące się nadmierną aktywnością ruchową, przejawiają się zmniejszeniem hamowania, jakie produkty wyjściowe (GPi i SNr) wywierają na wypustki wzgórzowo-korowe, które stają się bardziej aktywne..
Z drugiej strony zaburzeniom hipokinetycznym towarzyszy wzrost tego hamowania, ze zmniejszeniem aktywności wzgórzowo-korowej.
Jest to zaburzenie hiperkinetyczne charakteryzujące się mimowolnymi i spazmatycznymi przypadkowymi drżeniami kończyn i okolicy ustno-twarzowej, ruchami pląsawiczymi lub „tańczącymi”, które stopniowo narastają i unieruchamiają pacjenta, zaburzeniami mowy i postępującym rozwojem demencji.
Chorobie wcześnie towarzyszy zwyrodnienie neuronów prążkowia GABA (+ Encef.) Szlaku pośredniego.
Ponieważ te neurony nie hamują już neuronów GPe GABAergicznych, nadmiernie hamują jądro podwzgórza, które przestaje wzbudzać hamujące wyjścia (GPi i SNr), a projekcje wzgórzowo-korowe są odhamowane..
Polega na gwałtownych skurczach proksymalnych mięśni kończyn, które są wyrzucane z siłą w ruchach o dużej amplitudzie. Uszkodzenie w tym przypadku to degeneracja jądra podwzgórza, co skutkuje czymś podobnym do opisanego dla pląsawicy, chociaż nie przez hiperhamowanie, ale przez zniszczenie jądra podwzgórza.
Charakteryzuje się trudnością i opóźnieniem w inicjowaniu ruchów (akinezja), spowolnieniem ruchów (hipokinezą), twarzą bez wyrazu lub mimiką w masce, zmianą chodu ze zmniejszeniem towarzyszących ruchów kończyn podczas ruchu i drżeniem mimowolnym kończyny w spoczynku.
Uszkodzenie w tym przypadku polega na degeneracji układu nigrostriatalnego, które są dopaminergicznymi wypustkami, które rozpoczynają się od zwartego obszaru istoty czarnej (SNc) i łączą się z neuronami prążkowia, które dają początek ścieżkom pośrednim i bezpośrednim..
Tłumienie wzbudzenia, które włókna dopaminergiczne wywierają na komórki GABA (+ Sust. P) szlaku bezpośredniego, usuwa hamowanie, które wywierają one na wyloty GABAergiczne (GPi i SNr) w kierunku wzgórza, które jest teraz bardziej hamowane. intensywność. Jest to wtedy odhamowanie wyjść.
Z drugiej strony, tłumienie aktywności hamującej, którą dopamina wywiera na komórki GABA (+ Encef.) Szlaku pośredniego, uwalnia je i zwiększa hamowanie, które wywierają na komórki GABA GPe, co odhamowuje neurony komórki jądro podwzgórza, które następnie hiperaktywuje wyjścia.
Jak widać, końcowy rezultat wpływu degeneracji dopaminergicznej na dwa wewnętrzne szlaki, bezpośrednią i pośrednią, jest taki sam, niezależnie od tego, czy jest to odhamowanie, czy stymulacja wyjść GABA-ergicznych (GPi i SNr), które hamują jądra wzgórzowe i zmniejszają ich wyjście do kory, co wyjaśnia hipokinezę



Jeszcze bez komentarzy