Plik Cykliczne AMP lub 3 ', 5'-monofosforan adenozyny jest cyklicznym nukleotydem, który działa jako wtórny przekaźnik i jest częścią podstawowych elementów kontroli biochemicznej i komunikacji wewnątrzkomórkowej wielu żywych organizmów.
Jego istnienie wykazali prawie 70 lat temu Sutherland i Rall (1958), którzy opisali zjawisko gromadzenia się tego nukleotydu w komórkach wątroby w wyniku podania epinefryny (adrenaliny)..
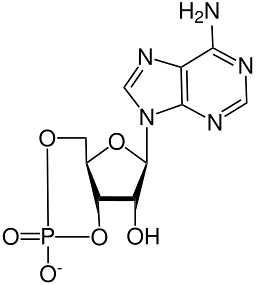
Od momentu odkrycia cykliczny AMP był kojarzony u ssaków z działaniem wielu hormonów, z wydzielaniem endokrynologicznym i zewnątrzwydzielniczym, między innymi z uwalnianiem neuroprzekaźników w synapsie i połączeniach nerwowo-mięśniowych..
Jego synteza jest katalizowana przez trzy rodziny białek związanych z błoną plazmatyczną, znane jako cyklaza adenylowa lub cyklaza adenylanowa, zdolne do wytwarzania cyklicznego związku z ATP i uwalniania pirofosforanu do komórki..
Z drugiej strony w jego degradacji pośredniczą enzymy z rodziny fosfodiesterazy, które są rozpuszczalnymi białkami występującymi głównie w cytozolu.
Te enzymy, a tym samym cykliczne AMP, znajdują się w niezwykle różnorodnych organizmach, tak prostych jak jednokomórkowe glony i wiele innych mikroorganizmów (bakterie i inne) oraz tak złożonych, jak zwierzęta wielokomórkowe o skomplikowanych szlakach sygnałowych..
Chociaż jej obecność w roślinach jest przedmiotem dyskusji, istnieją pewne dowody wskazujące na to, że niektóre gatunki roślin wykazują aktywność cyklazy adenylanowej, chociaż jej funkcja nie została w zadowalający sposób określona..
Indeks artykułów
Struktura chemiczna cyklicznego AMP została wyjaśniona za pomocą krystalografii rentgenowskiej i badań protonowego jądrowego rezonansu magnetycznego..
To jest termostabilna cykliczna cząsteczka („termostabilny ") i bardziej odporny na hydrolizę alkaliczną niż jego niecykliczny odpowiednik, AMP lub monofosforan adenozyny.
Podobnie jak wszystkie nukleotydy fosforanowe, cykliczny AMP ma grupę fosforanową przyłączoną do atomu węgla w pozycji 5 'cząsteczki rybozy, która z kolei jest przyłączona do heterocyklicznej zasady azotowej pierścienia poprzez węgiel w pozycji 1' i odpowiada adenina.
Grupa fosforanowa rybozy, w przeciwieństwie do niecyklicznych nukleotydów fosforanowych, jest skondensowana przeł poprzez wiązanie fosfodiestrowe z tlenami atomów węgla w pozycjach 3 'i 5' rybozy (3 ', 5'-trans-stopiony fosforan).
To połączenie ogranicza ruch pierścienia furanowego, który tworzy rybozę i zamyka grupę fosforanową w konformacji „krzesła”..
W porównaniu z niecyklicznymi nukleotydami, cykliczne AMP i inne pokrewne nukleotydy są mniejszymi cząsteczkami o niższej polarności, co jest ważnym czynnikiem ich różnicowania przez białka, które na nie odpowiadają..
Konformacja wiązania glikozydowego między rybozą a pierścieniem adeninowym ma pewną swobodę rotacji. Jest to również ważny parametr strukturalny umożliwiający odróżnienie go od innych nukleotydów (nie tylko tożsamość zasady azotowej).
Jako wtórny przekaźnik, cykliczny AMP uczestniczy w aktywacji wielu procesów sygnalizacyjnych (będących następstwem jego syntezy) lub w aktywacji różnych enzymów „w dół” kaskady sygnalizacyjnej, w której jest wytwarzany..
Uczestniczy w wątrobowej glikogenolizie oraz w uwalnianiu insuliny z trzustki, w uwalnianiu amylazy z gruczołów ślinowych oraz w działaniu estrogenu w macicy.
Ma uniwersalne funkcje w kontrolowaniu ekspresji genów i integracji wielu funkcji metabolicznych. Wiele cytokin wykorzystuje zarówno wapń, jak i cykliczne AMP do wykonywania swoich funkcji..
Wśród hormonów wykorzystujących cykliczne AMP w swoich procesach sygnalizacyjnych (poprzez zwiększanie lub zmniejszanie jego wewnątrzkomórkowego stężenia) można wymienić między innymi katecholaminy, glukagon, wazopresynę, parathormon, prostaglandyny, insulinę, melatoninę i adrenalinę..
Inną z jego wielu funkcji jest hamowanie wzrostu, różnicowania i proliferacji limfocytów T u ssaków, prawdopodobnie poprzez aktywację lub indukcję represora cytokin regulatorowych tych procesów w takich komórkach..
Cykliczny AMP i wytwarzające go cyklazy adenylanowe są również związane z funkcją wielu receptorów białkowych sprzężonych z białkiem G, które są związane z różnymi mechanizmami sygnalizacji i innymi ważnymi procesami komórkowymi..
U ssaków cykliczny AMP pełni funkcje w regulacji szlaku glikolitycznego i glukoneogennego poprzez hamowanie aktywności enzymu fosfofruktokinazy 2 (PFK-2), który katalizuje drugą reakcję glikolizy..
Mechanizm polega na udziale hormonu glukagonu w aktywacji wątrobowej cyklazy adenylanowej, co powoduje znaczny wzrost stężenia cyklicznego AMP..
Ten cykliczny AMP aktywuje zależną od cAMP kinazę białkową, która fosforyluje i hamuje aktywność fosfofruktokinazy PFK-2, który jest dwufunkcyjnym enzymem o aktywności bisfosfatazy fruktozowej..
Pierwszy przekaźnik (o zmiennej naturze chemicznej), który dociera do określonej komórki jako bodziec zewnętrzny, oddziałuje z enzymem cyklazy adenylanowej w błonie komórkowej, indukując produkcję cyklicznego AMP.
Wzrost stężenia cyklicznego AMP działa w aktywacji innych czynników (na ogół enzymatycznych), które pełnią dodatkowe funkcje m.in. w hamowaniu lub aktywacji procesów metabolicznych czy transkrypcji genów..
Jedną z głównych funkcji związanych z tą cząsteczką regulatorową jest aktywacja enzymów fosforylazy lub kinazy, które katalizują dodawanie lub usuwanie grup fosforylowych do innych białek i enzymów..
Normalnie wzbudzeniu komórki towarzyszy wzrost stężenia cyklicznego AMP, któremu towarzyszy wzrost transportu wapnia do komórki, która pełni funkcje w aktywacji enzymów cyklazy adenylowej wytwarzających cykliczne AMP..
Zarówno synteza, jak i przekazywanie wiadomości oraz degradacja cyklicznego AMP w komórkach są precyzyjnie regulowanymi procesami, które uczestniczą w utrzymaniu homeostazy organizmu.



Jeszcze bez komentarzy