Plik kardiolipina, znany również jako difosfatydyloglicerol, jest lipidem z rodziny glicerofosfolipidów oraz z grupy poliglicerofosfolipidów. Występuje w błonie mitochondrialnej organizmów eukariotycznych, w błonie plazmatycznej wielu bakterii, a także w niektórych archeonach.
Został odkryty przez Pangborna w 1942 roku na podstawie analizy lipidów błonowych tkanki serca bydła. Jego strukturę zaproponowano w 1956 roku, a synteza chemiczna miała miejsce około 10 lat później..
Niektórzy autorzy uważają, że jego obecność jest ograniczona do błon wytwarzających ATP, takich jak mitochondria u eukariotów, błony plazmatyczne bakterii i hydrogensomy (organelle podobne do mitochondriów) u niektórych protistów..
Fakt, że kardiolipina znajduje się w mitochondriach i błonie plazmatycznej bakterii, został wykorzystany do wzmocnienia podstaw teorii endosymbiotycznej, zgodnie z którą mitochondria powstały w komórkach progenitorowych eukariotów w wyniku fagocytozy bakterii, która następnie stała się komórką zależne i odwrotnie.
Jej szlak biosyntetyczny u zwierząt został opisany w latach 1970–1972, a później wykazano, że jest to ten sam szlak, który występuje u roślin, drożdży, grzybów i bezkręgowców. Nie jest to bardzo bogaty lipid, ale komórki wymagają go do prawidłowego funkcjonowania.
Znaczenie tego fosfolipidu dla mitochondriów, a tym samym dla metabolizmu komórkowego, jest oczywiste, gdy nieprawidłowe działanie związanych z nim szlaków metabolicznych powoduje u ludzi patologię znaną jako zespół Bartha (miopatia sercowo-szkieletowa).
Indeks artykułów
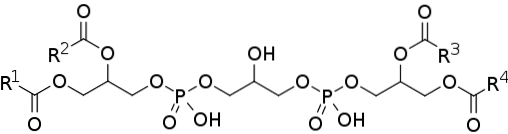
Kardiolipina lub difosfatydyloglicerol składa się z dwóch cząsteczek kwasu fosfatydowego (najprostszego fosfolipidu) połączonych ze sobą cząsteczką glicerolu.
Kwas fosfatydowy, jeden z powszechnych produktów pośrednich w szlakach biosyntezy innych fosfolipidów, składa się z cząsteczki 3-fosforanu glicerolu, do której dwa łańcuchy kwasów tłuszczowych są estryfikowane w pozycjach węgli 1 i 2, tak zwanych 3-fosforan diacyloglicerolu.
Dlatego kardiolipina składa się z trzech cząsteczek glicerolu: centralnej glicerolu, przyłączonej do grupy fosforanowej przy węglu 1, innej grupy fosforanowej przy węglu 3 i grupy hydroksylowej przy węglu 2; i dwie „boczne” glicerole.
Dwie „boczne” cząsteczki glicerolu są połączone z cząsteczką centralną „mostkami glicerolowymi” poprzez ich atomy węgla w pozycji 3. Na atomach węgla w pozycjach 1 i 2 estryfikowane są dwa łańcuchy kwasów tłuszczowych o zmiennej długości i nasyceniu..
Kardiolipina jest lipidem, który może tworzyć dwuwarstwy lub nie, w zależności od obecności lub braku dwuwartościowych kationów. Wiąże się to z faktem, że jest to cząsteczka symetryczna, co czyni ją ważną w błonach odpowiedzialnych za procesy transdukcji energii..
Podobnie jak inne lipidy z grupy poliglicerofosfolipidów, kardiolipina ma kilka grup hydroksylowych, które mogą służyć do wiązania kwasów tłuszczowych. Dlatego posiada wiele stereoizomerów pozycyjnych..
Różne badania wykazały, że kwasy tłuszczowe przyłączone do bocznych cząsteczek glicerolu kardiolipiny są zazwyczaj nienasycone, ale nie określono stopnia nienasycenia..
Zatem takie kwasy tłuszczowe mogą mieć długość 14-22 węgli i 0-6 wiązań podwójnych. To oraz fakt, że kardiolipina ma cztery połączone cząsteczki kwasów tłuszczowych, sugeruje, że może istnieć wiele zmiennych i kombinacji tego fosfolipidu..
Jak można się spodziewać, biosynteza kardiolipiny rozpoczyna się od syntezy kwasu fosfatydowego lub 3-fosforanu 1,2-diacyloglicerolu z 3-fosforanu glicerolu i kwasów tłuszczowych. Proces ten zachodzi w mitochondriach eukariotów oraz w błonie komórkowej bakterii..
Po utworzeniu kwas fosfatydowy reaguje z wysokoenergetyczną cząsteczką analogiczną do ATP: CTP. Następnie powstaje półprodukt, również o wysokiej energii, znany jako fosfatydylo-CMP. Aktywowana grupa fosfatydylowa jest przenoszona do grupy hydroksylowej w pozycji C1 centralnej cząsteczki glicerolu, która służy jako szkielet..
W wyniku tego procesu powstaje cząsteczka zwana fosfatydyloglicerofosforanem, która jest hydrolizowana z wytworzeniem fosfatydyloglicerolu. Otrzymuje inną aktywowaną grupę fosfatydylową z innej cząsteczki fosfatydylo-CMP, reakcję katalizowaną przez fosfatydylotransferazę znaną również jako syntaza kardiolipiny.
Enzym syntaza kardiolipiny znajduje się w wewnętrznej błonie mitochondrialnej i wydaje się tworzyć duży kompleks, przynajmniej w drożdżach. Jego gen ulega ekspresji w dużych ilościach w tkankach bogatych w mitochondria, takich jak serce, wątroba i mięśnie szkieletowe kręgowców..
Regulacja jego aktywności zależy w dużej mierze od tych samych czynników transkrypcyjnych i endokrynologicznych, które modulują biogenezę mitochondriów.
Po zsyntetyzowaniu w wewnętrznej błonie mitochondrialnej kardiolipina musi zostać przemieszczona w kierunku zewnętrznej błony mitochondrialnej, tak aby w błonie zachodziła seria procesów topologicznych i inne jej elementy strukturalne zostały przystosowane..
Zawartość kardiolipiny w bakteriach może być bardzo zmienna i zależy głównie od stanu fizjologicznego komórek: jest zwykle mniej obfita w wykładniczej fazie wzrostu, a obfitsza, gdy następuje jej redukcja (np. W fazie stacjonarnej).
Jej biosyntetyczny szlak może być wyzwalany przez różne stresujące bodźce, takie jak niedobory energii lub stres osmotyczny.
Aż do powstania fosfatydyloglicerolu proces u eukariontów i prokariotów jest równoważny, ale u prokariotów fosfatydyloglicerol otrzymuje przez transestryfikację grupę fosfatydylową z innej cząsteczki fosfatydyloglicerolu. Ta reakcja jest katalizowana przez enzym fosfolipazy typu D, znany również jako syntaza kardiolipiny..
Ta reakcja jest znana jako reakcja „transfosfatydylacji” (z angielskiego „transfosfatydylacja ”), gdzie jeden z fosfatydylogliceroli działa jako donor grupy fosfatydylowej, a drugi jako akceptor.
Fizyczne właściwości cząsteczek kardiolipiny najwyraźniej pozwalają na pewne interakcje, które odgrywają ważną rolę w strukturalnej organizacji błon, w których się znajdują..
Wśród tych funkcji jest między innymi rozróżnienie niektórych domen błonowych, interakcja lub „skrzyżowanie” z białkami transbłonowymi lub ich subdomenami..
Dzięki swoim właściwościom fizykochemicznym kardiolipina jest uznawana za lipid, który nie tworzy dwuwarstwy, ale którego funkcją może być stabilizacja i „przystosowanie” białek transbłonowych w dwuwarstwach lipidowych..
W szczególności jego właściwości elektryczne nadają mu funkcje w procesach przenoszenia protonów, które mają miejsce w mitochondriach.
Chociaż komórki mogą przetrwać bez tego fosfolipidu, niektóre badania wykazały, że jest on niezbędny do ich optymalnego funkcjonowania.



Jeszcze bez komentarzy