Plik guanina Jest to zasada azotowa, która służy do biosyntezy 5'-monofosforanu guanylanu i 5'-monofosforanu deoksyguanylanu. Obie substancje są odpowiednio częścią RNA i DNA, które przechowują informacje genetyczne komórek.
Kwas rybonukleinowy (RNA) i kwas dezoksyrybonukleinowy (DNA) składają się z nukleotydów, które składają się z zasady azotowej przyłączonej do cukru i grupy fosforanowej.
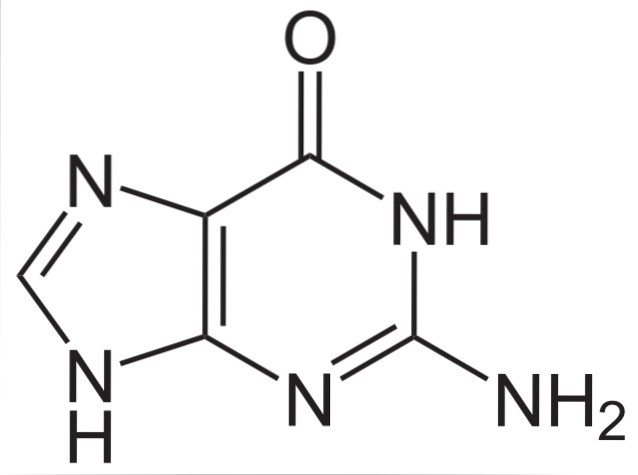
Guanina, oprócz tego, że jest częścią kwasów nukleinowych, w postaci monofosforanów, difosforanów i trifosforanów nukleozydów (GMP, GDP i GTP) uczestniczy w procesach takich jak metabolizm energetyczny, translacja sygnału wewnątrzkomórkowego, fizjologia fotoreceptorów i fuzja pęcherzyków.
Indeks artykułów
Struktura chemiczna guaniny (2-amino-6-hydroksypuryny) to heterocykliczny pierścień purynowy, składający się z układu dwóch połączonych ze sobą pierścieni: jeden to pirymidyna, a drugi to imidazol..
Pierścień heterocykliczny guaniny jest płaski i zawiera kilka sprzężonych wiązań podwójnych. Dodatkowo ma dwie formy tautomeryczne, keto i enol, pomiędzy grupami C-1 i N-6..
Cechy guaniny są następujące:
- Guanina jest substancją niepolarną. Jest nierozpuszczalny w wodzie, ale jest rozpuszczalny w stężonych roztworach mocnych kwasów lub zasad.
- Można go wyodrębnić jako białą substancję stałą o wzorze empirycznym C.5H.5N5O i ciężar cząsteczkowy 151,3 g / mol.
- Właściwość DNA do pochłaniania światła przy 260 nm wynika częściowo ze struktury chemicznej guaniny.
- W DNA guanina tworzy trzy wiązania wodorowe. Grupa karbonylowa C-6 jest akceptorem wiązań wodorowych, grupa N-1 i grupa aminowa C-2 są donorami wiązań wodorowych.
Z tego powodu rozerwanie wiązania między guaniną a cytozyną wymaga więcej energii niż w przypadku adeniny z tyminą, ponieważ ta ostatnia para jest połączona tylko dwoma wiązaniami wodorowymi..
- W komórce zawsze występuje jako część kwasów nukleinowych lub jako GMP, GDP i GTP, nigdy w postaci wolnej.
Cząsteczka guaniny, podobnie jak inne puryny, jest syntetyzowana de novo od 5-fosforybozylo-1-pirofosforanu (PRPP), poprzez reakcje katalizowane przez enzymy.
Pierwszy etap polega na dodaniu grupy aminowej z glutaminy do PRPP i utworzeniu 5-fosforybozyloaminy (PRA).
Następnie, w uporządkowanej kolejności, do PRA dodaje się glicynę, asparaginian, glutaminę, mrówczan i dwutlenek węgla. W ten sposób powstaje pośredni metabolit zwany 5'-monofosforanem inozyny (IMP)..
Podczas tego procesu darmowa energia jest wykorzystywana z hydrolizy ATP (5'-trifosforan adenozyny), z którego powstaje ADP (5'-difosforan adenozyny) i Pi (fosforan nieorganiczny)..
Utlenianie IMP zależy od NAD+ (dinukleotyd nikotynamidoadeninowy) wytwarza 5'-monofosforan ksantyny (XMP). Późniejsze dodanie grupy aminowej do XMP daje cząsteczkę guanylanu.
Regulacja biosyntezy guanylanu zachodzi na początku, gdy powstaje PRA, a na końcu, gdy zachodzi utlenianie IMP. Regulacja zachodzi poprzez negatywne sprzężenie zwrotne: nukleotyd GMP hamuje enzymy na obu etapach.
Podczas metabolicznej degradacji nukleotydów zawracane są zasady azotowe. GMP jest tworzony przez enzym fosforybozylotransferazę hipoksantynowo-guaninową, przenoszący grupę fosrybozylową z PRPP do guaniny.
Ponieważ guanina nie występuje w postaci wolnej, jej funkcje są powiązane z GMP, GDP i GTP. Niektóre z nich są wymienione poniżej:
- 5'-trifosforan guanozyny (GTP) działa jako rezerwuar darmowej energii. Grupa fosforanu gamma z GTP może zostać przeniesiona do adenozyny 5'-trifosforanu (ADP), z wytworzeniem ATP. Ta reakcja jest odwracalna i jest katalizowana przez nukleozydową kinazę difosforanową..
- GMP jest najbardziej stabilną formą nukleotydu zawierającą guaninę. Poprzez hydrolizę GMP tworzy cykliczny GMP (cGMP), który jest drugim przekaźnikiem podczas sygnalizacji wewnątrzkomórkowej w szlakach translacji. Na przykład w komórkach zapachowych fotoreceptorów i chemoreceptorów.
- CGMP uczestniczy w rozluźnieniu naczyń krwionośnych mięśni gładkich podczas biosyntezy tlenku azotu w komórkach śródbłonka.
- Hydroliza GTP gamma fosforanu służy jako wolne źródło energii do biosyntezy białek w rybosomach.
- Enzymy helicaza wymagają wolnej energii z hydrolizy GTP do oddzielenia podwójnej helisy DNA podczas replikacji i transkrypcji DNA.
- W neuronach hipokampu działanie bramkowanych napięciem kanałów sodowych jest regulowane przez hydrolizę GTP do GDP..
Wysoki poziom kwasu moczowego we krwi i moczu wiąże się z trzema różnymi defektami metabolicznymi, które zobaczymy poniżej..
Charakteryzuje się niedoborem HPRT (hipoksantyno-guaninofosforybozylotransferazy), enzymu ważnego dla recyklingu hipoksantyny i guaniny. W tym przypadku wzrasta poziom PRPP i nie powstają IMP i GMP, dwa ważne regulatory początkowego etapu syntezy puryn. Wszystko to sprzyja biosyntezie de novo puryn.
Powoduje to wzrost poziomów PRPP. Metabolit ten działa jako aktywator glutaminy PRPP-amidotransferazy, która odpowiada za syntezę 5-fosforybozyloaminy, zwiększając biosyntezę de novo puryny.
Jest to choroba związana z magazynowaniem glikogenu typu I. Pacjenci z tym zespołem mają wadliwą 6-fosfatazę glukozową. Powoduje to wzrost poziomu glukozo-6-fosforanu, który służy do syntezy rybozy-5-fosforanu poprzez pentozofosforan..
5-fosforan rybozy jest wyjściowym metabolitem biosyntezy PRPP. Podobnie jak w dwóch poprzednich przypadkach prowadzi to do wzrostu biosyntezy de novo puryn.
Zwiększony poziom kwasu moczowego we krwi i moczu powoduje objawy, które są powszechnie znane jako dna. W przypadku zespołu Lescha Nyhana pacjentom całkowicie brakuje aktywności enzymu HPRP, co prowadzi do manifestacji innych objawów, w tym paraliżu i upośledzenia umysłowego..
Gen HPRP znajduje się na chromosomie X. Dlatego mutacje w tym genie dotykają mężczyzn. Nie ma leczenia problemów neurologicznych. Objawy związane ze zwiększeniem stężenia kwasu moczowego leczy się allopurynolem.



Jeszcze bez komentarzy