
Plik dziedziczenie postmendlowskie Obejmuje wszystkie idee i / lub prace, które powstały po badaniach opublikowanych przez religijnego przyrodnika Gregorio Mendla w 1865 roku i które wyjaśniają zjawiska lub zachowania genetyczne inne niż opisane w tym dokumencie..
Gregorio Mendel jest uważany za ojca genetyki, ponieważ badając krzyżówki grochu, ponad 100 lat temu ustanowił pierwsze prawa dziedziczenia. Pierwsze prawo, zwane „prawem segregacji”, sugeruje, że geny mogą istnieć w różnych lub alternatywnych formach.

Te alternatywne formy nazywane są „allelami” i bezpośrednio wpływają na fenotypowe cechy gatunku. Allele mogą określać kolor włosów, kolor oczu, typ włosów itp..
W swoim pierwszym prawie Mendel zaproponował, że każda cecha fenotypowa przekazywana przez rodziców ich dzieciom jest kontrolowana przez dwa allele, każdy pochodzący od rodzica (jeden od „matki” i jeden od „ojca”).
Zgodnie z tym, gdy rodzice przekazują różne allele, potomstwo tworzy mieszaninę, w której jeden z alleli jest dominujący (ten, który jest wyrażony w fenotypie), a drugi jest recesywny (nie jest widoczny lub jest „utajony”) ”)..
Drugie prawo Mendla jest znane jako „Prawo niezależnego dziedziczenia”, ponieważ w swoich krzyżach Mendel zauważył, że wzór dziedziczenia, który charakteryzował allele definiujące cechę, nie wpływał na wzór dziedziczenia alleli, które definiowały inną cechę.
Indeks artykułów
Odkąd Gregor Mendel zaproponował swoje słynne prawa ponad 100 lat temu, wyjaśniono nowe mechanizmy wyjaśniające dziedziczenie postaci, z których wiele można powiedzieć, że są sprzeczne lub znacznie różnią się od tych opisanych przez Mendla..
Przykładem zjawisk opisywanych przez „nowoczesną” naukę, której postęp był możliwy tylko dzięki rozwojowi strategii eksperymentalnych i metod badawczych, jest m.in. dziedziczenie związane z płcią, niepełną dominacją i kodominacją..
Dziedziczenie związane z płcią ma związek z genami, które znajdują się tylko na chromosomach płci. U ludzi płeć jest określana przez chromosomy X i Y. Kobiety mają dwie kopie X (XX), a mężczyźni jedną z X i jedną z Y (XY)..
Geny znajdujące się na każdym z tych dwóch chromosomów to te, które przyczyniają się do tych cech, które są powiązane z płcią. Geny na chromosomie X są obecne u mężczyzn i kobiet, ale geny Y występują tylko u mężczyzn.
Dziedziczenie związane z płcią zostało zaobserwowane i opisane po raz pierwszy przez Williama Batesona, jednego z pierwszych naukowców, który „ponownie odkrył” prace Mendla i wykorzystał je do przeprowadzenia nowych eksperymentów opartych na jego wnioskach..
Bateson wykonał eksperymentalne krzyżówki kur z upierzeniami w różnych kolorach, niektóre z ciemnymi i gładkimi piórami, a inne z gładkimi piórami i pasiastym wzorem..
Poprzez te krzyże zauważył, że u potomstwa samce powstałe w wyniku zawsze miały upierzenie z pasiastym wzorem.
Dopiero w trzecim pokoleniu, gdy pasiastego samca skrzyżowano z kurą o ciemnych i gładkich piórach, uzyskano kilka kogutów o czarnym upierzeniu. Ta wynikająca z tego proporcja krzyży była sprzeczna z proporcjami proponowanymi przez Mendla.
Po pracy Batesona Thomas Henry Morgan znalazł logiczne wyjaśnienie dziwnych wyników uzyskanych przez Batesona, przypisując różnicę w zabarwieniu kurczaków faktowi, że wzór w paski był kodowany przez dominujący allel znajdujący się na chromosomie X..
U kur, w przeciwieństwie do ludzi, płeć heterogametyczna (XY) odpowiada samicom, natomiast płeć homogametyczna samcom (XX).
Allel badany przez Batesona i Morgana był zatem wyrażany przez wszystkich mężczyzn z upierzeniem w paski, ale gdy samiec odziedziczył dwa chromosomy X z allelem odpowiadającym czarnemu ubarwieniu, pojawiły się samce o ciemnym upierzeniu i bez pasiastego wzoru..
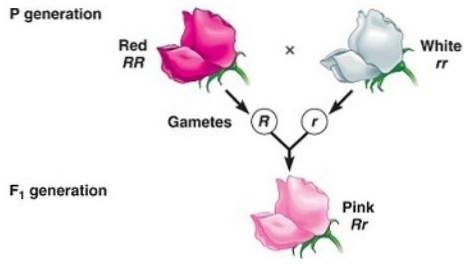
Dominacja niepełna występuje wtedy, gdy fenotyp obserwowany u potomstwa, który jest wynikiem skrzyżowania dwóch osobników homozygotycznych (jednego dominującego, a drugiego recesywnego) jest fenotypem „pośrednim” między dominującym a recesywnym..
Ma to związek z „niezdolnością” allelu „dominującego” do całkowitego maskowania ekspresji allelu recesywnego, dlatego powstały fenotyp odpowiada mieszaninie cech kodowanych przez oba allele, wielokrotnie opisywanej jako „rozcieńczony fenotyp ”Dominującej.
Na przykład w zakładach Dondiego w nocy (Mirabilis jalapa) typowy kolor płatków dzikiego kwiatu to czerwony. Kiedy krzyżuje się roślinę z czerwonymi płatkami z czystą linią o białych płatkach, uzyskuje się, że całe potomstwo ma różowe (różowe) płatki.
Kiedy krzyżuje się dwa dominujące osobniki homozygotyczne i obserwuje się trzeci fenotyp u potomstwa, który odpowiada dwóm cechom określonym przez dwa allele rodzicielskie, mówimy o kodominacji.
W tym przypadku, kontynuując przykład podobny do kwiatów z poprzedniego przykładu, „trzeci fenotyp” nie odpowiadałby różowym kwiatom, ale roślinie z białymi i czerwonymi kwiatami na tej samej łodydze, ponieważ te dwa allele „codominate”, I żadne z nich nie maskuje drugiego.
Jednak często kodominacja i niepełna dominacja są trudne do rozróżnienia fenotypowego, ponieważ oba tworzą fenotyp osobnika, który wydaje się wyrażać dwa różne allele..
Jednak genotypowo osoby, które prezentują allele kodominujące różnią się od tych, które wykazują niepełną dominację, ponieważ w jednym przypadku dwa dominujące allele są wyrażane w tej samej ilości, aw drugim proporcje są nierówne..
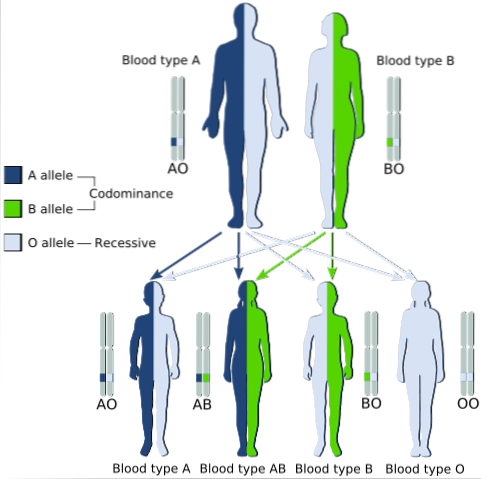
Klasycznym przykładem kodominacji jest typ grupy krwi u ludzi, ponieważ allele kodujące antygeny A i B w populacjach ludzkich są kodominujące.
Oznacza to, że kiedy allele A i B znajdują się razem w komórkach osobnika, ma on grupę krwi AB.
Są allelami recesywnymi dla niektórych postaci, które, gdy zostaną znalezione w formie homozygotycznej, są śmiertelne dla jednostki. Osoby, które prezentują te allele, nie rodzą się, ponieważ uniemożliwiają one normalne funkcjonowanie maszynerii molekularnej i uniemożliwiają translację genów.
Wiele miejsc w genomie osobnika oddziałuje z innymi częściami tego samego genomu, to znaczy specyficzna modyfikacja allelu w genomie może bezpośrednio warunkować ekspresję innych alleli w komórkach osobnika.
Te interakcje są trudne do określenia i można je podzielić na dwa różne typy: geny, które oddziałują na tym samym szlaku genetycznym i te, które oddziałują na różnych szlakach genetycznych..
Klasyczny i dobrze znany przykład genów, które oddziałują na różne ścieżki genetyczne, ma związek z genami, które określają kolor skóry węży kukurydzianych (Pantherophis guttatus), którego wspólnym kolorem jest powtarzający się wzór czarnych i pomarańczowych plamek.

Ten fenotyp jest wytwarzany przez dwa różne pigmenty, które podlegają ścisłej kontroli genetycznej. Gen określa kolor pomarańczowy i ma dwa allele: jeden O +, który koduje pigment, a drugi O-, który określa całkowity brak pomarańczowego pigmentu..
Inny inny gen określa obecność lub brak czarnego pigmentu, allele tego genu są znane jako B +, ten, który określa obecność czarnego pigmentu i B-, który określa brak pigmentu.
Oba geny nie są połączone i niezależnie określają kolor węży, to znaczy wąż z genotypem O + / B + jest normalnym wężem z pomarańczowym i czarnym fenotypem, ale każda zmienność genotypowa będzie skutkować innym fenotypem..
Klasycznym przykładem tych genów jest ten, który został opisany dla kwiatów „dzwonka” (z rodziny Campanulaceae), których „dziki” fenotyp odpowiada roślinom o niebieskich kwiatach..
Niebieski kolor kwiatów dzwonków nie jest określany przez gen, ale przez szlak metaboliczny, w którym kilka enzymów, każdy kodowany przez inny gen, uczestniczy w syntezie pigmentu..
Zatem istnieją fenotypy charakteryzujące się tym, że są wynikiem interakcji kilku genów, których produkty uczestniczą w tym samym szlaku metabolicznym i w których mutacja lub nieprawidłowe działanie któregokolwiek z nich daje fenotyp, w tym przypadku białych, odbarwionych kwiatów..



Jeszcze bez komentarzy