Plik cyklaza adenylanowa lub cyklaza adenylowa jest enzymem odpowiedzialnym za konwersję ATP, wysokoenergetycznej cząsteczki, do cyklicznego AMP, ważnej cząsteczki sygnałowej, która aktywuje różne zależne od cyklicznie białka AMP o ważnych funkcjach fizjologicznych.
Jego aktywność jest kontrolowana przez różne czynniki, takie jak na przykład skoordynowane działanie hormonów, neuroprzekaźników i innych cząsteczek regulatorowych o różnym charakterze (między innymi dwuwartościowe jony wapnia i białka G)..
Główne znaczenie tego enzymu polega na znaczeniu produktu reakcji, którą katalizuje, cyklicznego AMP, gdyż uczestniczy on w kontroli wielu zjawisk komórkowych związanych z metabolizmem i rozwojem, a także w odpowiedzi na różne bodźce zewnętrzne..
W naturze zarówno organizmy jednokomórkowe (stosunkowo proste), jak i duże i złożone zwierzęta wielokomórkowe używają cyklicznego AMP jako drugiego przekaźnika, a zatem enzymów, które go wytwarzają..
Badania filogenetyczne wykazały, że enzymy te pochodzą od wspólnego przodka przed rozdzieleniem eubakterii i eukariontów, co sugeruje, że cykliczny AMP miał różne funkcje, być może związane z produkcją ATP..
Można przyjąć takie stwierdzenie, ponieważ reakcja katalizowana przez cyklazę adenylanową jest łatwo odwracalna, co można zauważyć przy stałej równowagi dla syntezy ATP (K.eq ≈ 2,1 ± 0,2 10-9 Mdwa).
Indeks artykułów
Większość eukariotycznych enzymów cyklazy adenylanowej jest związanych z błoną plazmatyczną, ale w bakteriach i plemnikach ssaków występują one jako rozpuszczalne białka w cytozolu.
U drożdży i niektórych bakterii są białkami błony obwodowej, podczas gdy u niektórych gatunków ameb są cząsteczkami z pojedynczym segmentem transbłonowym.
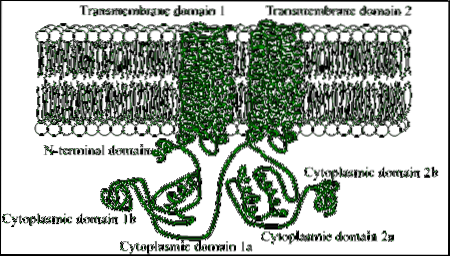
Są to białka złożone z dużych łańcuchów polipeptydowych (ponad 1000 reszt aminokwasowych), które przecinają błonę plazmatyczną 12 razy przez dwa regiony złożone z sześciu domen transbłonowych o konformacji alfa helisy..
Każdy region transbłonowy jest oddzielony dużą domeną cytozolową, która jest odpowiedzialna za aktywność katalityczną..
Wśród organizmów eukariotycznych są pewne konserwatywne motywy we fragmencie regionu aminoterminalnego tych enzymów, a także domena cytoplazmatyczna o masie około 40 kDa, ograniczona odcinkami hydrofobowymi..
Reakcja, którą katalizują te enzymy, to znaczy tworzenie wiązania diestrowego poprzez nukleofilowy atak grupy OH w pozycji 3 'na grupę fosforanową trifosforanu nukleozydu w pozycji 5', zależy od wspólnego motywu strukturalnego znanego jako domena Palm ”.
Ta domena „dłoni” składa się z motywu „βαβααβ„(„ Β ”oznacza arkusze złożone w β i„ α ”helisy alfa) i ma dwie niezmienne reszty kwasu asparaginowego, które koordynują dwa jony metali odpowiedzialne za katalizę, którymi mogą być dwuwartościowe jony magnezu lub magnezu. Cynk.
Wiele badań związanych z czwartorzędową strukturą tych enzymów ujawniło, że ich jednostka katalityczna istnieje jako dimer, którego tworzenie zależy od segmentów transbłonowych, które łączą się w siateczce endoplazmatycznej podczas tworzenia białka..
Stwierdzono, że podobnie jak wiele integralnych białek błonowych, takich jak białka G, te z kotwicami fosfatydyloinozytolu i wiele innych, cyklazy adenylowe znajdują się w specjalnych regionach błonowych lub mikro-domenach znanych jako „tratwy lipidowe” (z angielskiego „Tratwa lipidowa”).
Te domeny błonowe mogą mieć średnicę do setek nanometrów i składają się głównie z cholesterolu i sfingolipidów z długimi i przeważnie nasyconymi łańcuchami kwasów tłuszczowych, co czyni je mniej płynnymi i umożliwia umieszczanie transbłonowych segmentów różnych białek.
Cyklazy adenylanowe zostały również znalezione w podregionach tratw lipidowych znanych jako „caveolae” (z angielskiego „caveolae ”), które są raczej wgłębieniami błony bogatej w cholesterol i związane z nim białko zwane kaweoliną.
W naturze istnieją trzy dobrze zdefiniowane klasy cyklazy adenylanowej i dwie, które są obecnie przedmiotem dyskusji..
- Klasa I: są obecne w wielu bakteriach Gram-ujemnych, takich jak E coli, na przykład, gdy cykliczny produkt reakcji AMP pełni funkcję liganda dla czynników transkrypcyjnych odpowiedzialnych za regulację operonów katabolicznych.
- Klasa II: występuje w niektórych patogenach z rodzajów bakterii, takich jak Bakcyl lub Bordetella, gdzie służą jako toksyny zewnątrzkomórkowe. Są to białka aktywowane przez kalmodulinę żywiciela (nieobecne w bakteriach).
- Klasa III: są znane jako klasa „uniwersalna” i są filogenetycznie spokrewnione z cyklazami guanylanowymi, które pełnią podobne funkcje. Występują zarówno u prokariontów, jak i eukariontów, gdzie są regulowane różnymi ścieżkami.
U ssaków co najmniej dziewięć typów tych enzymów zostało sklonowanych i opisanych, kodowanych przez dziewięć niezależnych genów i należących do cyklazy adenylowej klasy III..
Mają one złożone struktury i topologie błon, a także zduplikowane domeny katalityczne, które są dla nich charakterystyczne..
W przypadku ssaków nomenklatura stosowana w odniesieniu do izoform odpowiada literom AC (od cyklaza adenylanowa) i liczbę od 1 do 9 (AC1 - AC9). Doniesiono również o dwóch wariantach enzymu AC8.
Izoformy obecne u tych zwierząt są homologiczne pod względem sekwencji pierwotnej struktury ich miejsc katalitycznych i struktury trójwymiarowej. Włączenie jednego z tych enzymów do każdego „typu” jest głównie związane z mechanizmami regulacyjnymi, które działają na każdą izoformę.
Mają wzorce ekspresji, które często są specyficzne dla tkanki. Wszystkie izoformy można znaleźć w mózgu, chociaż niektóre są ograniczone do określonych obszarów ośrodkowego układu nerwowego.
Główną funkcją enzymów należących do rodziny cyklaz adenylanowych jest przekształcanie ATP w cykliczne AMP iw tym celu katalizują one tworzenie wewnątrzcząsteczkowego wiązania diestrowego 3'-5 '(reakcja podobna do tej katalizowanej przez polimerazy DNA). cząsteczki pirofosforanu.
U ssaków różne warianty, które można osiągnąć, były związane z proliferacją komórek, uzależnieniem od etanolu, plastycznością synaps, uzależnieniem od leków, rytmem okołodobowym, stymulacją węchową, uczeniem się i pamięcią..
Niektórzy autorzy zasugerowali, że cyklazy adenylanowe mogą pełnić dodatkową funkcję jako cząsteczki transporterów lub, co jest tym samym, białka kanałowe i transportery jonowe..
Jednak te hipotezy zostały przetestowane tylko w oparciu o układ lub topologię segmentów transbłonowych tych enzymów, które mają pewne homologie lub podobieństwa strukturalne (ale nie sekwencję) z pewnymi kanałami transportującymi jony..
Zarówno cykliczny AMP, jak i PPi (pirofosforan), które są produktami reakcji, pełnią funkcje na poziomie komórkowym; ale ich znaczenie zależy od organizmu, w którym się znajdują.
Ogromne zróżnicowanie strukturalne cyklaz adenylowych wskazuje na dużą podatność na wiele form regulacji, co pozwala im na integrację z wieloma różnymi szlakami sygnalizacji komórkowej..
Aktywność katalityczna niektórych z tych enzymów zależy od alfa ketokwasów, podczas gdy inne mają znacznie bardziej złożone mechanizmy regulacyjne, które obejmują podjednostki regulatorowe (poprzez stymulację lub hamowanie), które zależą na przykład od wapnia i innych ogólnie rozpuszczalnych czynników, a także innych białka.
Wiele cyklaz adenylanowych jest regulowanych negatywnie przez podjednostki niektórych białek G (hamują ich funkcję), podczas gdy inne wywierają bardziej aktywujący wpływ.


Jeszcze bez komentarzy