Plik chromatyna Jest to kompleks złożony z DNA i białek, unikalny w organizmach eukariotycznych. Pod względem proporcji zawiera prawie dwa razy więcej białek niż materiał genetyczny. Najważniejszymi białkami tego kompleksu są histony - małe, dodatnio naładowane białka, które wiążą się z DNA poprzez oddziaływania elektrostatyczne. Dodatkowo chromatyna zawiera ponad tysiąc innych białek niż histony..
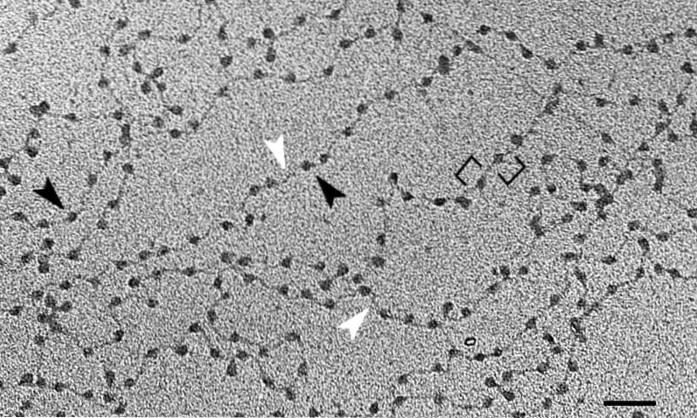
Podstawową jednostką chromatyny jest nukleosom, który składa się z połączenia histonów i DNA. Taki układ przypomina koraliki na naszyjniku. Po przejściu przez wszystkie wyższe poziomy organizacji DNA dotrzemy do chromosomów.
Struktura chromatyny jest ściśle związana z kontrolą ekspresji genów. Istnieją dwa główne typy: euchromatyna i heterochromatyna.
Euchromatyna charakteryzuje się niskim stopniem zagęszczenia, co przekłada się na wysoki poziom transkrypcji. Natomiast heterochromatyna jest nieaktywna transkrypcyjnie ze względu na wysoki stopień zagęszczenia..
Strukturalnie istnieją pewne epigenetyczne znamiona histonów typowe dla obu typów chromatyny. Podczas gdy euchromatyna jest związana z acetylacją, heterochromatyna jest związana ze spadkiem grup chemicznych.
Istnieją pewne regiony heterochromatyny z funkcjami strukturalnymi, takie jak telomery i centromery.
Indeks artykułów
Badania materiału genetycznego i jego organizacji strukturalnej rozpoczęły się w 1831 roku, kiedy badacz Robert Brown opisał jądro. Jednym z bezpośrednich pytań związanych z tym odkryciem było zapytanie o biologiczną i chemiczną naturę tej struktury..
Kwestie te zaczęto wyjaśniać między 1870 a 1900 rokiem dzięki eksperymentom Friedricha Mieschera, który wprowadził słowo nukleina. Jednak Walther Flemming modyfikuje termin i używa chromatyna odnosić się do substancji jądrowej.
Z biegiem czasu zaczynasz głębiej rozumieć materiał genetyczny i jego właściwości. Dopiero w 1908 roku włoski badacz Pasquale Baccarini zauważył, że chromatyna nie jest jednorodna i zdołała uwidocznić małe ciała wewnątrz jądra..
Rodzaje chromatyny - euchromatyna i heterochromatyna - zostały po raz pierwszy zaproponowane przez Emila Heitza w 1928 r. Aby ustalić tę klasyfikację, Heitz oparł się na zastosowaniu barwników.
W 1974 roku biolog Roger Kornberg zaproponował model organizacji materiału genetycznego w strukturach znanych jako nukleosomy, hipoteza potwierdzona empirycznie przez eksperymenty Markusa Noll..
Chromatyna to nukleoproteina utworzona przez połączenie materiału genetycznego - DNA - z heterogenicznym zestawem białek. To skojarzenie jest bardzo dynamiczne i uzyskuje złożoną trójwymiarową konformację, która pozwala mu pełnić funkcje regulacyjne i strukturalne..
Jednym z najważniejszych białek chromatyny są histony, które występują w prawie takiej samej proporcji jak DNA.
Histony to podstawowe białka, niezwykle konserwowane w całej ewolucyjnej historii istot organicznych - to znaczy, że nasze histony nie różnią się zbytnio w porównaniu z histonami innych ssaków, nawet innego zwierzęcia bardziej odległego filogenetycznie.
Ładunek histonów jest dodatni, więc mogą oddziaływać poprzez siły elektrostatyczne z ujemnym ładunkiem szkieletu fosforanowego obecnego w DNA. Istnieje pięć typów histonów, a mianowicie: H1, H2A, H2B, H3 i H4.
Istnieje również szereg białek o innym charakterze niż histony, które biorą udział w zagęszczaniu DNA..
Podstawową jednostką chromatyny są nukleosomy - powtarzalne struktury zbudowane z DNA i histonów, konformacji występującej w całym materiale genetycznym..
Podwójna helisa DNA jest zwinięta w kompleks ośmiu histonów, znanych jako oktamer histonu. Cząsteczka zwija się w przybliżeniu w dwóch zwojach, po których następuje krótki region (między 20 a 60 parami zasad), który oddziela nukleosomy od siebie..
Aby zrozumieć tę organizację, musimy wziąć pod uwagę, że cząsteczka DNA jest niezwykle długa (około 2 metry) i musi być zwinięta w uporządkowany sposób, aby osadzić się w jądrze (którego średnica wynosi od 3 do 10 µm). Ponadto należy go udostępnić do replikacji i transkrypcji..
Cel ten osiąga się przy różnych poziomach zagęszczenia DNA, z których pierwszym są wyżej wymienione nukleosomy. Przypominają one koraliki na naszyjniku z pereł. Około 150 par zasad DNA jest zranionych w domowych „zliczeniach”.
W bakteriach nie ma prawdziwych historii. W przeciwieństwie do tego istnieje szereg białek przypominających histony i przypuszcza się, że przyczyniają się one do pakowania bakteryjnego DNA..
Organizacja chromatyny nie jest ograniczona na poziomie nukleosomów. Ta asocjacja białek i DNA jest zgrupowana w grubszej strukturze około 30 nm - ze względu na tę grubość nazywana jest poziomem „włókna 30 nm”..
Chromatyna zorganizowana w grubości 30 nm jest z kolei zorganizowana w postaci pętli, które rozciągają się w swego rodzaju rusztowanie o charakterze białkowym (nie histony).
Jest to obecnie stosowany model, chociaż można oczekiwać istnienia bardziej złożonych mechanizmów zagęszczania. Ostateczna organizacja składa się z chromosomu.
Zagęszczenie i organizacja materiału genetycznego ma kluczowe znaczenie dla wielu funkcji biologicznych. Różne schorzenia wiązały się z błędami w budowie chromatyny, w tym między innymi talasemią alfa sprzężoną z chromosomem X, zespołem Rubinsteina-Taybiego, zespołem Coffina-Lowry'ego, zespołem Retta..
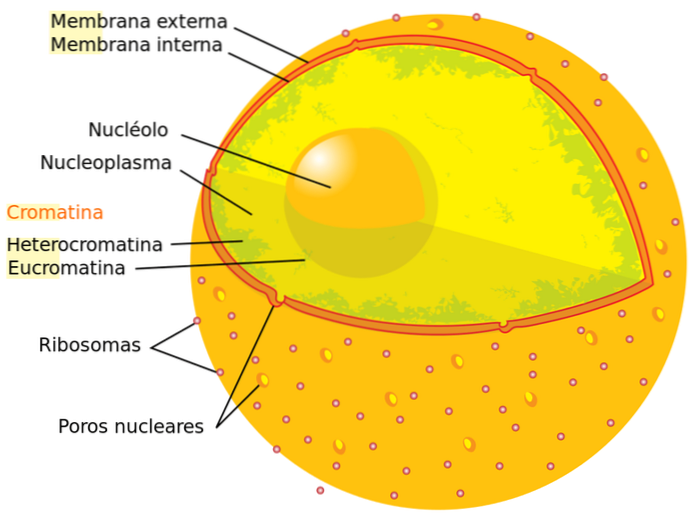
Istnieją dwa rodzaje chromatyny w komórce, ujawniające się po zastosowaniu barwników: euchromatyna („prawdziwa” chromatyna) i heterochromatyna. W pierwszym przypadku zabarwienie jest słabo widoczne, w drugim barwienie jest intensywne.
Ta strukturalna organizacja DNA jest unikalna dla organizmów eukariotycznych i ma kluczowe znaczenie dla zachowania chromosomów i regulacji ekspresji genów..
Jeśli ocenimy proporcje obu typów chromatyny w komórce znajdującej się w interfazie, stwierdzimy, że około 90% chromatyny to euchromatyna, a pozostałe 10% to heterochromatyna. Poniżej szczegółowo opiszemy każdy typ:
Główna różnica, jaka istnieje między oboma typami chromatyny, jest związana ze stopniem zagęszczenia lub „upakowania” cząsteczki na określonych etapach podziału komórki..
Chociaż na styku materiał genetyczny wydaje się być losowo rozproszony, tak nie jest.
Na tym etapie istnieje znacząca organizacja, w której wewnątrz jądra można zobaczyć zróżnicowaną partycję materiału chromosomowego.
DNA chromosomów nie splata się z nicią DNA innych chromosomów i pozostaje w określonych regionach zwanych terytoriami chromosomowymi. Wydaje się, że ta organizacja przyczynia się do ekspresji genów.
Heterochromatyna jest silnie skondensowana, przez co jest niedostępna dla mechanizmu transkrypcyjnego - więc nie podlega transkrypcji. Ponadto jest ubogi pod względem liczby posiadanych genów..
Niektóre regiony heterochromatyny są trwałe w całej linii komórkowej - tj., na zawsze zachowa się jak heterochromatyna. Ten typ heterochromatyny jest znany jako konstytutywny. Przykładem tego są skondensowane regiony chromosomów zwane centromerami i telomerami..
W przeciwieństwie do tego, istnieją porcje heterochromatyny, które mogą zmieniać poziom zagęszczenia w odpowiedzi na zmiany wzorców rozwoju lub zmienne środowiskowe..
Dzięki nowym badaniom wizja ta jest przeformułowywana i obecnie istnieją dowody na to, że konstytutywna heterochromatyna jest również dynamiczna i zdolna do reagowania na bodźce..
Jednym z czynników determinujących budowę chromatyny są chemiczne modyfikacje histonów. W przypadku chromatyny nieaktywnej transkrypcyjnie wykazują one hipoacetylowane histony.
Zmniejszenie liczby grup acetylowych wiąże się z wyciszaniem genów, ponieważ nie maskuje dodatniego ładunku lizyn, umożliwiając silną interakcję elektrostatyczną między DNA i histonami..
Kolejną marką epigenetyczną jest metylacja. Ponieważ jednak dodanie grupy metylowej nie modyfikuje ładunku białka, jego konsekwencja (aktywacja lub dezaktywacja genów) nie jest tak oczywista i będzie zależeć od regionu histonu, w którym znajduje się znak..
Stwierdzono empirycznie, że metylacja H3K4me3 i H3K36me3 jest związana z aktywacją genów, a H3K9me3 i H3K27me3.
Na przykładzie konstytutywnej heterochromatyny wymieniamy centromer. Ten region chromosomowy pełni rolę strukturalną i bierze udział w ruchu chromosomów zarówno podczas mitotycznych, jak i mejotycznych wydarzeń podziału komórek..
W przeciwieństwie do heterochromatyny, euchromatyna jest mniej zwartą cząsteczką, więc maszyneria transkrypcyjna ma łatwy dostęp (szczególnie enzym polimerazy RNA) i może być aktywnie wyrażana na szlakach genetycznych.
Struktura nukleosomu aktywnej transkrypcyjnie chromatyny jest scharakteryzowana przez acetylowane histony. Obecność monometylowanych lizyn jest również związana z aktywacją genów.
Dodanie grupy acetylowej do tych reszt histonów lizyny neutralizuje dodatni ładunek wspomnianego aminokwasu. Bezpośrednią konsekwencją tej zmiany jest redukcja oddziaływań elektrostatycznych między histonem i DNA, co prowadzi do powstania luźniejszej chromatyny.
Ta modyfikacja strukturalna umożliwia interakcje materiału genetycznego z maszynerią transkrypcyjną, która charakteryzuje się wyjątkowo dużą masą..
Euchromatyna obejmuje wszystkie aktywne geny, do których można dotrzeć za pomocą mechanizmu enzymatycznego związanego z transkrypcją. Dlatego funkcje są tak szerokie, jak funkcje zaangażowanych genów..


Jeszcze bez komentarzy