Plik włókna Purkinjego Komórki serca stanowią ostatni etap systemu, który automatycznie i wielokrotnie wytwarza wzbudzenie elektryczne wymagane do mechanicznej aktywności komór. Koncentruje się na kierowaniu pobudzenia do miocytów komorowych, tak aby wytwarzały skurcz (skurcz).
System, do którego należą te włókna, składa się z węzła zatokowo-przedsionkowego (SA), z którego pochodzi wzbudzenie; wiązki międzywęzłowe, które docierają do węzła przedsionkowo-komorowego (AV); węzeł przedsionkowo-komorowy, w którym przewodzenie elektryczne jest nieco opóźnione; wiązka Jego, z jego prawą i lewą gałęzią oraz system włókien Purkinjego.
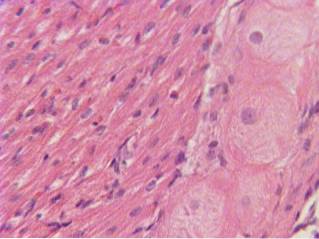
Włókna te zostały nazwane na cześć Johna Evangelisty Purkinjego, czeskiego anatoma i fizjologa, który po raz pierwszy opisał je w 1839 roku. Nie należy ich mylić z komórkami Purkinjego, odkrytymi przez tego samego autora na poziomie kory móżdżku i zaangażowanymi w kontrolę ruchu..
Indeks artykułów
Podobnie jak pozostałe elementy układu pobudzenia i przewodzenia w sercu, komórki tworzące system włókien Purkinjego to komórki mięśniowe lub miocyty serca, które utraciły swoją kurczliwą strukturę i wyspecjalizowały się w przewodzeniu wzbudzenia elektrycznego..
Jego składniki łączą końce gałęzi wiązki His i początek sekwencji miocytów komorowych, między którymi przewodzą pobudzenie elektryczne pochodzące z węzła zatokowo-przedsionkowego, tworząc rozproszoną sieć rozmieszczoną w całym wsierdziu pokrywającym komory ..
Posiadają cechy, które odróżniają je od innych elementów systemu: są dłuższymi i grubszymi włóknami (40 μm) nawet od włókien kurczliwych komorowych i mają największą prędkość przewodzenia: 4 m / s; W porównaniu z następującymi po nim 1,5 m / s, włókna wiązki Hisa.
Ta wysoka prędkość przewodzenia wynika, oprócz dużej średnicy, z faktu, że w miejscach ich styku z interkalowanymi dyskami występuje duża gęstość połączeń szczelinowych (skrzyżowania szczelinowe), które umożliwiają łatwy przepływ prądów jonowych między nimi i szybkie przenoszenie wzbudzenia.
Ze względu na tę dużą prędkość przewodzenia i rozproszoną dystrybucję włókien Purkinjego, pobudzenie dociera prawie jednocześnie do kurczliwego mięśnia sercowego obu komór, wymagając zaledwie 0,03 s (30 ms) do pełnej aktywacji całego mięśnia sercowego..
Komórki układu Purkinjego są komórkami pobudliwymi, które w spoczynku wykazują różnicę potencjałów od -90 do -95 mV między obiema powierzchniami błony, która oddziela jej wnętrze od otaczającego płynu zewnątrzkomórkowego, przy czym jej wnętrze jest ujemne w stosunku do zewnętrznej.
Po pobudzeniu komórki te reagują depolaryzacją zwaną potencjałem czynnościowym (AP), podczas której potencjał błonowy szybko staje się mniej ujemny i może zostać odwrócony, osiągając chwilowo dodatnią wartość do +30 mV (dodatnia wewnątrz).
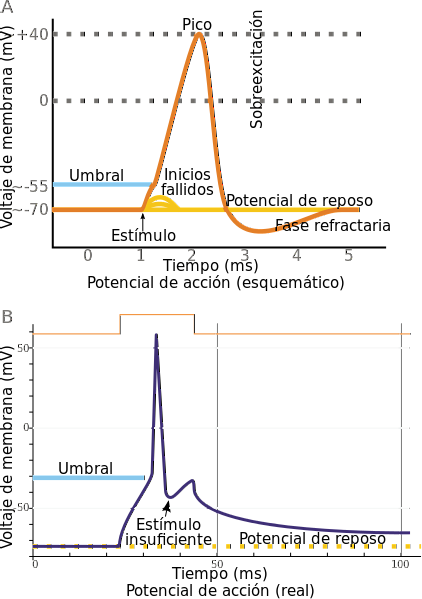
W zależności od szybkości, z jaką zachodzi ta depolaryzacja, różne typy komórek pobudliwych serca zostały zaliczone do jednej z dwóch kategorii: włókna szybko reagujące lub włókna o wolnej odpowiedzi. Włókna Purkinje należą do tej drugiej kategorii.
Fizjologicznym bodźcem dla włókien Purkinjego do wytworzenia potencjału czynnościowego jest depolaryzujący prąd jonowy, pochodzący z elementów komórkowych, które znajdują się wcześniej w sekwencji przewodzenia, i dociera do nich przez szczeliny łączące je z tymi elementami..
W potencjale czynnościowym włókna Purkinjego wyróżnia się kilka faz: nagła depolaryzacja (faza 0) do +30 mV, szybka repolaryzacja do 0 mV (faza 1), długotrwała depolaryzacja około 0 mV (faza 2 lub plateau) i szybka repolaryzacja (faza 3) prowadząca z powrotem do potencjału spoczynkowego (faza 4).
Zdarzenia te są wynikiem aktywacji i / lub dezaktywacji prądów jonowych, które modyfikują równowagę ładunków między wnętrzem i zewnętrzem ogniw. Prądy, które z kolei wynikają ze zmian przepuszczalności poszczególnych kanałów dla różnych jonów i są oznaczone literą I, po której następuje indeks dolny, który je identyfikuje.
Prądy wejściowe jonów dodatnich lub prądy wyjściowe jonów ujemnych są zgodnie z konwencją uważane za ujemne i powodują depolaryzację, natomiast prądy wyjściowe jonów dodatnich lub wyjściowych jonów ujemnych są prądami dodatnimi i sprzyjają wewnętrznej polaryzacji lub negatywie komórki..
Faza 0 występuje, gdy początkowa depolaryzacja, która służy jako bodziec, sprowadza potencjał błony do poziomu (progu) między -75 a -65 mV, a następnie otwierają się zależne od napięcia kanały sodowe (Na +), które umożliwiają wejście Na + (prąd Ina ) jak w lawinie, zwiększając potencjał do około +30 mV.
Faza 1 rozpoczyna się na końcu fazy 0, kiedy kanały Na + ponownie zamykają się i depolaryzacja ustaje, wytwarzając prądy przejściowe (Ito1 i Ito2) wyjścia K + i wejścia Cl-, które powodują szybką repolaryzację do poziomu 0 mV.
Faza 2 jest to „plateau” o długim czasie trwania (300 ms). Wynika to z otwarcia wolnych kanałów wapniowych i wytworzenia dopływu Ca ++, który wraz z remanentnym dopływem Na + utrzymuje stosunkowo wysoki potencjał (0 mV) i przeciwdziała prądom repolaryzacji K + (IKr i IKs), które zaczęły się pojawiać.
Na faza 3 Prądy Ca ++ i Na + są zminimalizowane, a prądy repolaryzacji na wylocie K + stają się bardzo wyraźne. Ta rosnąca wydajność K + sprowadza potencjał błony do początkowego poziomu spoczynkowego od -90 do -95 mV, na którym pozostaje (faza 4) dopóki cykl się nie powtórzy.
- Poziom spoczynkowy: -90 do -95 mV.
- Maksymalny poziom depolaryzacji (przeregulowanie): + 30 mV.
- Amplituda potencjału czynnościowego: 120 mV.
- Czas trwania potencjału czynnościowego: od 300 do 500 ms.
- Szybkość depolaryzacji: 500-700 V / s.
- Poziom progowy wyzwalania potencjału czynnościowego: między -75 a -65 mV.
- Prędkość jazdy: 3-4 m / s.
Wolno reagujące włókna mięśnia sercowego obejmują komórki węzłów zatokowo-przedsionkowych i przedsionkowo-komorowych, które w stanie spoczynku (faza 4) ulegają powolnej depolaryzacji (rozkurczowo-prepotencjalnej), która podnosi potencjał błony do poziomu progowego i potencjału czynnościowego. uruchamiane automatycznie.
Ta właściwość jest bardziej rozwinięta, to znaczy depolaryzacja zachodzi szybciej w węźle zatokowo-przedsionkowym, który służy jako rozrusznik serca i charakteryzuje się częstością od 60 do 80 uderzeń / min. Jeśli to się nie powiedzie, węzeł przedsionkowo-komorowy może przejąć polecenie, ale z mniejszą częstotliwością między 60 a 40 uderzeń / min.
Włókna Purkinjego, gdy nie są wzbudzane przez normalny układ przewodzący, mogą również podlegać temu samemu procesowi powolnej depolaryzacji, która doprowadza ich potencjał błonowy do poziomu progowego i automatycznie wypala potencjały czynnościowe.
W przypadku, gdy normalne wzbudzenie węzła zatokowo-przedsionkowego i wtórnego węzła przedsionkowo-komorowego zawodzi lub przejście pobudzenia do komór jest zablokowane, niektóre włókna układu Purkinjego zaczynają się rozładowywać samodzielnie i utrzymują rytmiczna komora aktywacji, ale z mniejszą szybkością (25-40 uderzeń / min).

Jeszcze bez komentarzy