
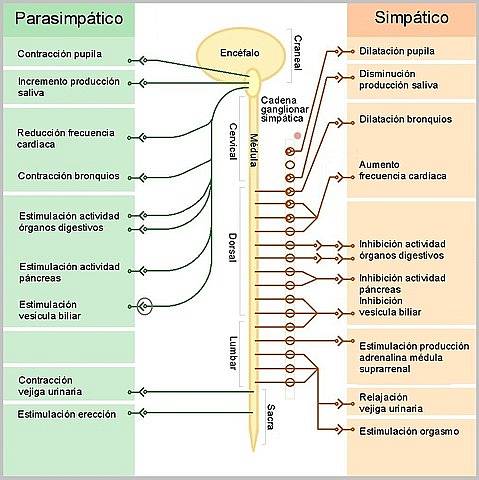
Plik unerwienie serca jest zorganizowany w unerwienie współczulne i przywspółczulne. Jak każdy inny narząd, serce otrzymuje unerwienie, którego włókna są klasyfikowane jako należące do autonomicznego układu nerwowego (ANS), jednego z dwóch oddziałów obwodowego układu nerwowego i odpowiedzialne za pośredniczenie w wrażliwości i kontrolowanie czynności trzewnej organizmu.
Pomimo tego, że jest narządem mięśni poprzecznie prążkowanych, bardzo podobnym do mięśni szkieletowych, serce nie podlega unerwieniu z innego podziału układu obwodowego, który pośredniczy w wrażliwości somatycznej i aktywności mięśni, która powoduje przemieszczenia stawów.
Każdy proces skurczu mięśni szkieletowych wymaga pobudzenia wywołanego przez somatyczne włókno nerwu ruchowego. Serce ze swej strony nie musi być wzbudzane przez nic zewnętrznego dla siebie, ponieważ ma zdolność spontanicznego generowania własnych podnieceń..
Tak więc jedną z wyjątkowych cech unerwienia autonomicznego serca jest fakt, że nie jest ono wyznacznikiem czynności skurczowej serca, która może trwać po odnerwieniu, ale raczej pełni jego funkcję modulującą..
Indeks artykułów
Eferentna lub motoryczna część autonomicznego układu nerwowego jest podzielona na dwa komponenty: współczulny i przywspółczulny, składające się ze szlaków łączących neurony w ośrodkowym układzie nerwowym z trzewnymi komórkami efektorowymi ciała, na które wywierają działanie antagonistyczne..
Każdy z tych szlaków jest łańcuchem dwóch neuronów:
Współczulne komórki preganglionowe przeznaczone dla serca pochodzą z konglomeratów komórkowych zlokalizowanych w rogach bocznych rdzenia kręgowego, w odcinku piersiowym T1-T5. Konglomeraty komórek, które razem tworzą „rdzeniowy ośrodek współczulny kardioakceleratora”.
Jej aksony reprezentują włókna przedwojenne, które są skierowane do współczulnego łańcucha zwojowego; szczególnie do górnych, środkowych i dolnych zwojów szyjnych, gdzie łączą się z neuronami post zwojowymi, których aksony są rozmieszczone z górnym, środkowym i dolnym nerwem serca.
Spośród tych trzech nerwów, środkowy wydaje się być tym, który wywiera największy wpływ na funkcje serca, ponieważ górny jest przeznaczony dla dużych tętnic u podstawy serca, a dolny wydaje się przewodzić informacje czuciowe lub aferentne..
Kolejnym szczegółem organizacji sercowego unerwienia współczulnego jest to, że prawe włókna współczulne wydają się kończyć głównie w węźle zatokowo-przedsionkowym, podczas gdy lewe wpływają na węzeł przedsionkowo-komorowy, układ przewodzenia i kurczliwy mięsień sercowy..
Współczulny układ nerwowy wpływa pozytywnie na wszystkie funkcje serca, zwiększając częstość akcji serca (chronotropizm +), siłę skurczu (inotropizm +), przewodzenie wzbudzenia (dromotropizm +) i prędkość relaksacji (lusotropizm +).
Wszystkie te działania wywoływane są poprzez uwalnianie noradrenaliny (NA) na poziomie współczulnych zakończeń postganglionicznych na komórkach węzłów sercowych, na układzie przewodzenia lub na kurczliwych miocytach przedsionkowo-komorowych..
Działanie noradrenaliny jest wyzwalane, gdy ten neuroprzekaźnik wiąże się z receptorami adrenergicznymi typu β1 znajdującymi się na błonach komórek serca i sprzężonymi z białkiem Gs. Jest to białko z trzema podjednostkami (αsβγ), które w stanie nieaktywnym wiąże GDP ze swoją podjednostką αs.
Oddziaływanie receptora norepinefryny-β1 powoduje, że podjednostka α uwalnia swój GDP i zamienia go na GTP; W ten sposób oddziela się od składnika βγ i aktywuje błonowy enzym cyklazę adenylową, który wytwarza cykliczny monofosforan adenozyny (cAMP) jako drugi przekaźnik, który aktywuje kinazę białkową A (PKA).
Aktywność fosforylująca PKA jest ostatecznie odpowiedzialna za wszystkie działania stymulujące, które włókna współczulne wywierają na serce, i obejmuje fosforylację kanałów Ca ++, troponiny I i fosfolamban.
Działanie na kanały Ca ++ sprzyja wzrostowi tętna, siły skurczu i szybkości przewodzenia. Wpływ na troponinę I i fosfolamban przyspiesza proces relaksacji mięśnia sercowego.
Fosforylacja troponiny I powoduje, że białko to przyspiesza proces uwalniania Ca ++ z troponiny C, dzięki czemu relaksacja następuje szybciej. Fosfolamban naturalnie hamuje pompę, która ponownie wprowadza Ca ++ do siateczki sarkoplazmatycznej w celu zakończenia skurczu, hamowanie, które jest zmniejszane, gdy jest fosforylowane.
Przywspółczulne unerwienie serca przebiega przez nerw błędny, a jego składowe mają układ bineuronalny podobny do łańcucha współczulnego, z neuronami przedwojowymi, których ciała znajdują się w grzbietowym jądrze motorycznym nerwu błędnego w opuszce czwarta komora..
Ze względu na zmniejszające się skutki czynności serca, które te neurony wywierają na serce, nazwano je zbiorczo „ośrodkiem hamującym krążenie opuszkowe”. Jego włókna oddzielają się od pnia błędnego w szyi, a następnie łączą się z włóknami współczulnymi serca, tworząc splot..
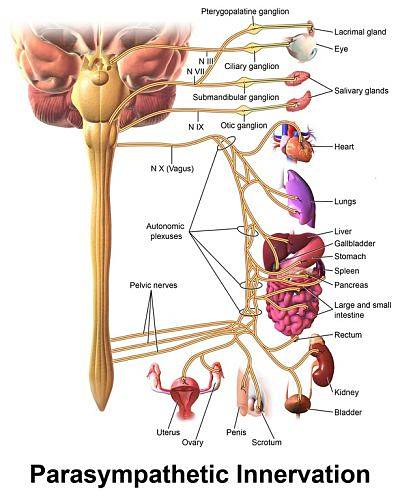
Zwoje przywspółczulne znajdują się w pobliżu serca, a włókna pozwojowe po prawej stronie kończą się głównie w węźle zatokowo-przedsionkowym, naturalnym rozruszniku serca, a lewe w węźle przedsionkowo-komorowym i w miocytach kurczliwych przedsionka.
Aktywność przywspółczulna skierowana na serce ma negatywny wpływ na niektóre funkcje serca, takie jak zmniejszenie częstotliwości (inotropizm -), szybkości przewodzenia w węźle AV (dromotropizm -) i zmniejszenie siły skurczu przedsionków (słuchawka inotropiczna - ).
Niewielkie lub wręcz brak unerwienia układu przywspółczulnego mięśnia sercowego komorowego wyklucza negatywny wpływ inotropowy tego autonomicznego podziału na siłę skurczową tego mięśnia.
Wspomniane powyżej działania nerwu błędnego na serce wywoływane są przez uwalnianie acetylocholiny (ACh) na poziomie przywspółczulnych zakończeń pozwojowych na komórkach węzłów serca i miocytach kurczliwych przedsionka..
Działanie acetylocholiny jest wyzwalane, gdy wiąże się ona z muskarynowymi receptorami cholinergicznymi typu M2 zlokalizowanymi na błonach wspomnianych komórek i sprzężonymi z białkiem Gi. Ma trzy podjednostki (αiβγ), a gdy jest nieaktywny, ma GDP dołączone do jego podjednostki αi..
Oddziaływanie acetylocholiny-receptora M2 uwalnia podjednostkę αi. To hamuje cyklazę adenylową, wytwarzane jest mniej cAMP i zmniejsza się aktywność PKA i fosforylacji kanałów Ca ++, efekty przeciwne do efektów NA uwalnianych przez układ współczulny. Składnik βγ aktywuje prąd K + (IKACh).
Zmniejszenie fosforylacji kanałów Ca ++ zmniejsza prąd depolaryzujący tego jonu, podczas gdy pojawienie się prądu IKACh wprowadza prąd hiperpolaryzujący, który przeciwdziała spontanicznej depolaryzacji, która wytwarza potencjały czynnościowe (AP) w komórkach guzkowych..
Zmniejszenie depolaryzującego prądu Ca ++ w połączeniu ze wzrostem hiperpolaryzującego prądu K + spowalnia spontaniczny proces depolaryzacji, który automatycznie sprowadza potencjał błony do poziomu progowego, przy którym wyzwalany jest potencjał czynnościowy..
Efekt ten może być tak duży, że intensywna stymulacja nerwu błędnego może zatrzymać serce, z powodu zaniku potencjałów czynnościowych komórek rozrusznika lub z powodu całkowitego zablokowania węzła przedsionkowo-komorowego, który nie pozwala na przejście potencjałów działanie od prawego przedsionka do komór.
Zarówno układ współczulny, jak i przywspółczulny są zawsze aktywne, wywierając trwałe działanie tonizujące na serce, tak że spoczynkowe funkcje serca są wynikiem spontanicznej aktywności serca modulowanej tonicznie przez te dwa antagonistyczne wpływy..
Ton przywspółczulny jest wyższy niż ton współczulny, co wynika z faktu, że gdy serce jest „odnerwione” chirurgicznie lub farmakologicznie, przyspiesza, zwiększając częstość akcji serca.
Zwiększone zapotrzebowanie metaboliczne organizmu wymaga zwiększenia czynności serca, które jest osiągane automatycznie poprzez zwiększenie działania układu współczulnego na serce i zmniejszenie działania przywspółczulnego. Stopień maksymalnego odpoczynku osiąga się przy odwrotnych działaniach.
Modulacja ośrodków kardioakceleratora i kardioinhibitorów, wspomnianych źródeł unerwienia autonomicznego serca, zależy od aktywności wyższych ośrodków nerwowych zlokalizowanych w pniu mózgu, podwzgórzu i korze mózgowej..

Jeszcze bez komentarzy