Plik mikrofilamenty lub filamenty aktynowe, są jednym z trzech podstawowych składników cytoszkieletu komórek eukariotycznych (mikrofilamentów, mikrotubul i włókienek pośrednich) i składają się z małych włókien białka zwanego aktyną (polimery aktyny).
U eukariontów geny kodujące mikrofilamenty aktyny są silnie konserwowane we wszystkich organizmach, dlatego często są wykorzystywane jako markery molekularne do badań różnego rodzaju.

Mikrofilamenty są rozmieszczone w całym cytozolu, ale są szczególnie liczne w regionie leżącym pod błoną komórkową, gdzie tworzą złożoną sieć i łączą się z innymi specjalnymi białkami, tworząc cytoszkielet..
Sieci mikrowłókien w cytoplazmie komórek ssaków są kodowane przez dwa z sześciu genów opisanych dla aktyny, które są zaangażowane w dynamikę mikrofilamentów, a nawet są bardzo ważne podczas różnicowania komórek macierzystych.
Wielu autorów zgadza się, że mikrofilamenty są najbardziej zróżnicowanymi, wszechstronnymi i ważnymi białkami w cytoszkieletie większości komórek eukariotycznych i należy pamiętać, że nie występują one w mikroorganizmach prokariotycznych..
Z drugiej strony w tego typu komórkach znajdują się włókna, które są homologiczne do mikrofilamentów, ale które składają się z innego białka: białka MreB..
Obecnie uważa się, że gen kodujący to białko może być genem przodków eukariotycznej aktyny. Jednak homologia sekwencji aminokwasów tworzących białko MreB wynosi tylko 15% w odniesieniu do sekwencji aktyny.
Ponieważ są one podstawową częścią cytoszkieletu, każdy defekt fenotypowy zarówno w mikrotubulach, jak i we włóknach pośrednich i mikrofilamentach aktyny (cytoszkielet) może powodować różne patologie komórkowe i ogólnoustrojowe..
Indeks artykułów
Mikrowłókna zbudowane są z monomerów białek z rodziny aktyn, które są białkami kurczliwymi bardzo obficie w komórkach eukariotycznych, ponieważ uczestniczą również w skurczach mięśni..
Włókna te mają średnicę od 5 do 7 nm, więc są również znane jako cienkie włókna i składają się z dwóch form aktyny: postaci kulistej (aktyna G) i postaci nitkowatej (aktyna F)..
Białka, które uczestniczą w cytoszkieletu, są znane jako γ i β aktyny, podczas gdy te, które uczestniczą w skurczu, to zwykle α aktyny..
Udział aktyny kulistej i aktyny nitkowatej w cytozolu zależy od potrzeb komórkowych, ponieważ mikrowłókna są strukturami wysoce zmiennymi i wszechstronnymi, które stale rosną i skracają się w wyniku polimeryzacji i depolimeryzacji..
Aktyna G to małe białko globularne, składające się z prawie 400 aminokwasów i o masie cząsteczkowej około 43 kDa.
Monomery G-aktyny, które tworzą mikrowłókna, są ułożone w postaci spiralnej nici, ponieważ każdy podlega skręceniu, gdy jest powiązany z następnym.
Aktyna G wiąże się z jedną cząsteczką Ca2 + i inną ATP, co stabilizuje jej postać kulistą; podczas gdy F-aktyna jest otrzymywana po hydrolizie końcowego fosforanu cząsteczki ATP do G-aktyny, co przyczynia się do polimeryzacji.
Włókna aktynowe mogą być organizowane w „wiązki” lub „sieci”, które pełnią różne funkcje w komórkach. Wiązki tworzą równoległe struktury połączone dość sztywnymi mostkami krzyżowymi.
Z drugiej strony sieci to luźniejsze struktury, przypominające trójwymiarowe siatki o właściwościach półstałych żeli..
Istnieje wiele białek, które są związane z filamentami aktyny lub mikrofilamentami i są znane jako ABP (z angielskiego białka wiążące aktynę), które mają do tego określone witryny.
Wiele z tych białek umożliwia interakcję mikrofilamentów z dwoma pozostałymi składnikami cytoszkieletu: mikrotubulami i włóknami pośrednimi, a także z innymi składnikami wewnętrznej powierzchni błony komórkowej..
Inne białka, z którymi oddziałują mikrofilamenty, obejmują blaszki jądrowe i spektynę (w krwinkach czerwonych).
Ponieważ globularne monomery aktyny zawsze wiążą się w ten sam sposób, zorientowane w tym samym kierunku, mikrofilamenty mają określoną biegunowość, z dwoma końcami: jednym „więcej” i jednym „mniej”..
Biegunowość tych włókien jest bardzo ważna, ponieważ rosną one znacznie szybciej na ich dodatnim końcu, do którego dodawane są nowe monomery G-aktyny..

Pierwszą rzeczą, jaka zachodzi podczas polimeryzacji włókien aktyny, jest proces zwany „zarodkowaniem”, który polega na asocjacji trzech monomerów białka.
Nowe monomery są dodawane do tego trimeru na obu końcach, tak że włókno rośnie. Monomery G-aktyny są zdolne do hydrolizy ATP przy każdym wiązaniu, co ma wpływ na szybkość polimeryzacji, ponieważ reszty aktyna-ATP dysocjują z większą trudnością niż reszty aktyny-ADP..
ATP nie jest konieczny do polimeryzacji, a szczególna rola jego hydrolizy nie została jeszcze wyjaśniona..
Niektórzy autorzy uważają, że skoro zdarzenia polimeryzacji aktyny są szybko odwracalne, ATP związane z tymi procesami może stanowić do 40% całkowitego obrotu komórkowego tej cząsteczki energii..
Zarówno polimeryzacja filamentów aktyny, jak i ich depolimeryzacja są procesami silnie regulowanymi przez szereg specyficznych białek, które są odpowiedzialne za przebudowę włókien..
Przykładem białek regulujących depolimeryzację jest czynnik depolimeryzacji aktyny, kofilina. Inne białko, profilina, pełni odwrotną funkcję, ponieważ stymuluje asocjację monomerów (poprzez stymulację wymiany ADP na ATP).
Mikrofilamenty oddziałują z filamentami miozyny, które są związane z białkami transbłonowymi, które mają domenę w cytozolu, a drugą na zewnątrz komórki, uczestnicząc w ten sposób w procesach mobilności komórki.
Te mikrowłókna związane z błoną plazmatyczną pośredniczą w różnych odpowiedziach komórkowych na różne klasy bodźców. Na przykład adhezja komórek w tkankach nabłonkowych jest napędzana przez białka transbłonowe zwane kadherynami, które oddziałują z mikrowłókienkami w celu rekrutacji czynników odpowiedzi..
Filamenty aktyny oddziałują z włóknami pośrednimi, powodując przekazywanie bodźców zewnątrzkomórkowych do kluczowych miejsc, takich jak rybosomy i chromosomy w jądrze..
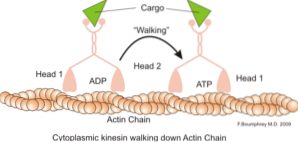
Klasyczną i dobrze zbadaną funkcją mikrowłókien jest ich zdolność do tworzenia „mostów”, „szyn” lub „autostrad” dla ruchu białka motorycznego miozyny I, która jest zdolna do ładowania pęcherzyków transportowych z organelli do plazmatycznej błony w wydzielinie. ścieżki.
Mikrofilamenty oddziałują również z miozyną II, tworząc kurczliwy pierścień, który tworzy się podczas cytokinezy, a dokładnie podczas ostatniego etapu podziału komórki, w którym cytozol jest oddzielany od komórek macierzystych i potomnych..
Ogólnie rzecz biorąc, mikrofilamenty F-aktyny modulują rozmieszczenie niektórych organelli, takich jak kompleks Golgiego, retikulum endoplazmatyczne i mitochondria. Ponadto biorą udział w przestrzennym pozycjonowaniu mRNA, dzięki czemu są odczytywane przez rybosomy..
Cały zestaw komórkowych mikrowłókien, zwłaszcza tych, które są blisko spokrewnione z błoną plazmatyczną, bierze udział w tworzeniu falujących błon komórek, które mają stały aktywny ruch.
Są również zaangażowani w tworzenie mikrokosmków i innych typowych guzków na powierzchni wielu komórek..
Mikrowłókna biorą udział w procesie wydzielania żółci w hepatocytach (komórkach wątroby), a także w ruchach perystaltycznych (skoordynowany skurcz) kanałów wątrobowych.
Przyczyniają się do różnicowania domen błony komórkowej dzięki ich powiązaniu z różnymi elementami cytozolowymi oraz kontroli, jaką wywierają nad topografią tych elementów wewnątrzkomórkowych..
Niewiele jest chorób związanych z pierwotnymi defektami w strukturze lub z białkami regulatorowymi i enzymami w syntezie mikrowłókien, mimo że są one bezpośrednio zaangażowane w wiele funkcji..
Niski wskaźnik chorób i wad rozwojowych w pierwotnej strukturze mikrofilamentów wynika z faktu, że na ogół istnieje wiele genów kodujących zarówno aktynę, jak i jej białka regulatorowe, zjawisko znane jako „nadmiarowość genetyczna”.
Jedną z najczęściej badanych patologii jest witryfikacja oocytów na ich cytoszkieletu, gdzie obserwuje się przerwanie sieci mikrowłókien korowych, a także depolimeryzację i dezorganizację mikrotubul wrzeciona mitotycznego..
Ogólnie rzecz biorąc, zeszklenie to powoduje dyspersję chromosomów, ponieważ prowadzi do zaburzenia zagęszczenia całej chromatyny.
Komórki, które mają większą organizację i udział mikrofilamentów w swoim cytoszkieletu to komórki mięśnia prążkowanego, dlatego większość patologii wiąże się z nieprawidłowym działaniem aparatu kurczliwego.
Wadliwe lub nietypowe mikrofilamenty są również związane z chorobą kości zwaną chorobą Pageta..


Jeszcze bez komentarzy