Plik podwójne zapłodnienie to proces zachodzący w roślinach, w wyniku którego jedno z jąder gametycznych ziarna pyłku (gametofit męski) zapładnia żeńską komórkę rozrodczą (oosferę), a inne zapładnia inną komórkę, która będzie inna, jeśli będzie to okrytozalążkowa lub gnetal.
U okrytozalążkowych drugie jądro gametofitu łączy się z dwoma polarnymi jądrami obecnymi w centralnej komórce worka pyłkowego, aby później rozwinąć się w bielmie. Z drugiej strony u gnetali drugie jądro męskiego gametofitu łączy się z jądrem kanału brzusznego, tworząc drugi zarodek..
Botanicy początkowo uważali, że podwójne zapłodnienie jest wyłącznym zjawiskiem u roślin okrytozalążkowych, jednak proces ten został później opisany również dla roślin z grupy Gnetal..
Indeks artykułów
Botanik pochodzenia rosyjskiego Sergey Gavrilovich Nawashin zajmujący się roślinami okrytozalążkowymi tego gatunku Lilium martagon Y Fritillaria tenella jako pierwszy zaobserwował proces podwójnego zapłodnienia. Zjawisko to zostało później wykazane przez słynnego polsko-niemieckiego botanika Eduarda Strasburgera.
Po tym odkryciu różni botanicy donosili o nieprawidłowych zdarzeniach podwójnego zapłodnienia w różnych grupach nagonasiennych (Tuja, Abies, Pseudotsuga, m.in). Produkty drugiego zapłodnienia mogą zdegenerować się, wytworzyć wolne jądra lub też spowodować powstanie dodatkowych zarodków.
Później wykazano, że podwójne zapłodnienie było normalnym zdarzeniem u roślin z grupy Gnetal, ale u nich, w przeciwieństwie do okrytozalążkowych, drugie zapłodnienie zawsze daje dodatkowe zarodki, a nie bielmo..
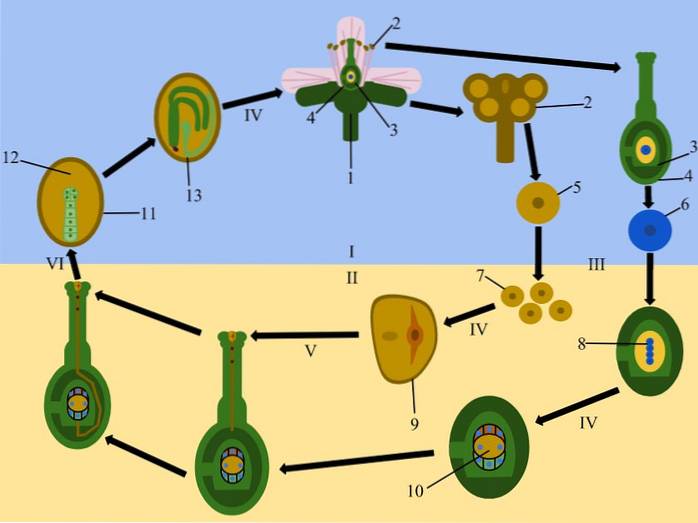
U większości roślin okrytozalążkowych, komórka prekursorowa megaspory (megasporocyte) wytwarza w wyniku mejozy cztery haploidalne megaspory, z których tylko jedna rozwija się do produkcji megagametofitu, podczas gdy inne ulegają degeneracji.
Megagametofit wytwarza osiem jąder, z których dwa (jądra polarne) migrują w kierunku centralnej strefy megagametofitu lub worka zarodkowego, tworząc w ten sposób komórkę dwujądrową..
Reszta jąder jest ułożona w grupach po trzy na obrzeżach, z których jedna utworzy ovocell, dwa sąsiednie utworzą synergie, a pozostałe trzy, znajdujące się na przeciwległym końcu, utworzą antypody..
Ze swojej strony męski gametofit (ziarno pyłku) wytwarza trzy jądra; dwa plemniki i jeden wegetatywny. Kiedy ziarno pyłku wchodzi w kontakt ze znamieniem, kiełkuje i wytwarza łagiewkę pyłkową, która rośnie wraz z pyleniem, przyciągana przez substancje wytwarzane przez synergetyków..
Następnie dwa jądra plemników migrują przez łagiewkę pyłkową, aby przeprowadzić podwójne zapłodnienie. Jedno z jąder plemników połączy się z jądrem komórki jajowej i utworzy zygotę, z której powstanie zarodek, podczas gdy drugie połączy się z dwoma jądrami komórki centralnej, dając początek bielmie.
Bielmo jest tkanką triploidalną powstałą w wyniku połączenia centralnej komórki worka zarodkowego (dwujądrowe) z jednym z dwóch jąder plemników ziaren pyłku. Komórki bielma są bogate w granulki skrobi osadzone w macierzy białkowej, a ich funkcją jest dostarczanie materiału odżywczego do rozwijającego się zarodka.
Wśród naukowców nie ma zgody co do ewolucyjnego pochodzenia bielma roślin okrytozalążkowych. Niektórzy autorzy twierdzą, że jest to dodatkowy zarodek, którego rozwój został przekształcony w tkankę odżywczą z korzyścią dla drugiego zarodka.
Inni autorzy argumentują, że bielmo nie jest produktem rozmnażania płciowego, ale jest fazą wegetatywną rozwoju woreczka zarodkowego, tak jak ma to miejsce u nagonasiennych. Obie hipotezy mają swoich przeciwników i obrońców.
Jedyne rośliny nagozalążkowe, u których udowodniono podwójne nawożenie, należą do rodzajów Gnetum Y Efedra (Gnatales). Jednak w żadnym z nich bielmo nie powstaje w wyniku tego podwójnego zapłodnienia..
W co najmniej dwóch gatunkach Efedra samice tworzą się w archegonii w obrębie gametofitów monosporycznych. Ze swojej strony plemnik zawiera cztery jądra, z których dwa są rozrodcze.
Te jądra plemników są uwalniane w megagametoficie i jeden z nich łączy się z jądrem komórki jajowej, podczas gdy drugi łączy się z jądrem kanału brzusznego..
Rezultatem są dwa zygoty, które rozwiną się w genetycznie identyczne żywotne zarodki, ponieważ dwa jądra żeńskie powstały w monosporycznym gametoficie, podczas gdy dwa jądra plemników z tej samej łagiewki pyłkowej są również genetycznie identyczne..
W każdym żeńskim gametoficie mogą powstać dwie lub więcej archegonii, co skutkuje wieloma zdarzeniami podwójnego zapłodnienia występującymi jednocześnie w gametoficie..
Podwójne nawożenie w Gnetum przedstawia istotne różnice w porównaniu z podwójnym zapłodnieniem, które występuje w Efedra. Na Gnetum, w przeciwieństwie do Efedra, samice gametofity są tetrasporyczne i nie są monosporyczne.
Inną różnicą jest to, że w Gnetum nie powstają ani archegonia, ani predestynowane komórki jajowe. Z powodu utraty komórek jajowych, liczne żeńskie jądra gametofityczne są zdolne do zapłodnienia; w ten sposób dwa jądra plemników z łagiewki pyłkowej mogą zapłodnić dowolne dwa jądra żeńskie.
Jak w Efedra, proces podwójnego zapłodnienia w Gnetum wyprodukuje dwie zdolne do życia zygoty, ale w tej zygoty nie będą genetycznie identyczne ze względu na tetrasporyczną naturę żeńskiego gametofitu. Na Gnetum przy wystarczającej ilości ziaren pyłku mogą również wystąpić wielokrotne przypadki podwójnego zapłodnienia.

Odkrycie podwójnego zapłodnienia w Gnetum Y Efedra popiera hipotezę ewolucyjną, zgodnie z którą ten proces powstał u wspólnego przodka Gnetales i okrytozalążkowych, dla którego byłby to synapomorfia (wspólny charakter pochodny), która pozwoliłaby na zgrupowanie ich w klad antofitów (monofiletyczne).
Synapomorfie to cechy pochodne, które są wspólne dla dwóch lub więcej gatunków lub taksonów, a zatem mogą wykazywać pewien stopień pokrewieństwa. Charakter przodków (plezjomorficzny) w tym przypadku byłby prostym zapłodnieniem.
W ten sposób Gnetales może być grupą podstawową w kladzie antofitów, w której podwójne zapłodnienie powoduje powstanie dwóch żywotnych zygot, podczas gdy pojawienie się bielma jako produktu podwójnego zapłodnienia byłoby unikalną synapomorfią w okrytozalążkowych..



Jeszcze bez komentarzy